quote:Originally posted by BrandonP: There is of course the recent argument that H. sapiens may be a "composite" species that came about after populations from different regions of the continent intermixed. Some people have taken to calling it "African multiregionalism":
Unfortunately most of the article is behind a paywall, but the caption summarizes it thus:
quote:Several new discoveries suggest that our species didnt arise from a single point in space. Instead, the entire continent was our cradle.
Indeed I've always questioned how anyone-- geneticist or paleo-anthropologist-- could pinpoint an exact area or locality in Africa where the species originated.
Here is the 2018 study that a lot of this is based on: 'Did Our Species Evolve in Subdivided Populations across Africa, and Why Does It Matter?' [I couldn't post the url link but it's an issue of cell.com]
The study suggests that there were periods of isolation between various populations (bottleneck events) followed by periods of extensive geneflow. The former caused by arid conditions while the latter wet conditions. Thus pinpointing an exact area in Africa as the one specific spot where the species arose is virtually impossible. Indeed, there is the suggestion that Sapiens could have been the result of various subspecies comingling through the process I just outlined.
Djehuti Member # 6698
posted
The same year the above study was published Scientific American wrote an article on it which I happen to grab an issue of.
The Last Hominin Standing: Why Did Homo Sapiens Alone Survive to the Modern Era? By Kate Wong At the dawning of Homo sapiens, our ancestors were born into a world we would find utterly surreal. Its not so much that the climate and sea levels or the plants and the animals were different, although of course they wereits that there were other kinds of humans alive at the same time. For most of H. sapiens existence, in fact, multiple human species walked the earth. In Africa, where our species got its start, large-brained Homo heidelbergensis and small-brained Homo naledi also roamed. In Asia, there was Homo erectus, a mysterious group dubbed the Denisovans and, later, Homo floresiensisa hobbitlike creature, tiny but for its large feet. The stocky, heavy-browed Neanderthals, for their part, ruled Europe and western Asia. And there were probably even more forms, as yet undiscovered. By around 40,000 years ago, based on current evidence, H. sapiens found itself all alone, the only remaining member of what was once an incredibly diverse family of bipedal primates, together known as hominins. (In this article, the terms human and hominin both refer to H. sapiens and its extinct relatives.) How did our kind come to be the last human standing? Until a few years ago, scientists favored a simple explanation: H. sapiens arose relatively recently, in more or less its current form, in a single region of Africa and spread out from there into the rest of the Old World, supplanting the Neanderthals and other archaic human species it encountered along the way. There was no appreciable interspecies fraternizing, just wholesale replacement of the old guards by the cleverer newcomer, whose ascendancy seemed inevitable. Yet mounting evidence from fossil and archaeological discoveries, as well as DNA analyses, has experts increasingly rethinking that scenario. It now looks as though H. Sapiens originated far earlier than previously thought, possibly in locations across Africa instead of a single region, and that some of its distinguishing traitsincluding aspects of the brainevolved piecemeal. Moreover, it has become abundantly clear that H. sapiens actually did mingle with other human species it encountered and that interbreeding with them may have been a crucial factor in our success. Together these findings paint a far more complex picture of our origins than many researchers had envisionedone that privileges the role of dumb luck over destiny in the success of our kind.
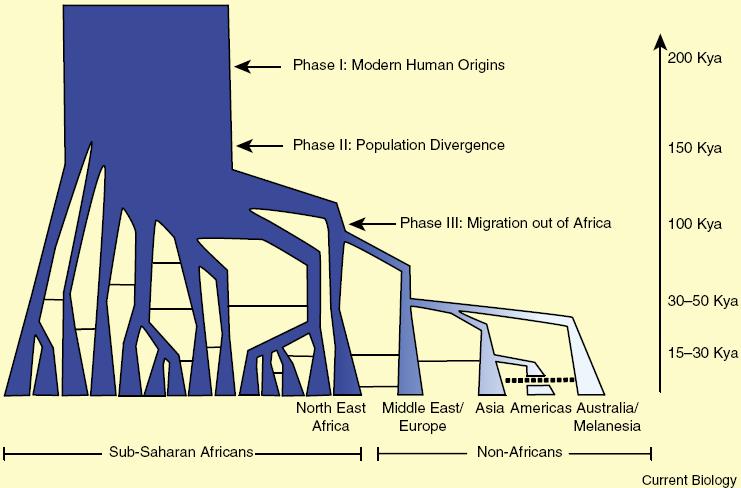
THEORY UNDER THREAT Debate about the origin of our species has traditionally focused on two competing models. One one side was the Recent African Origin hypothesis, championed by paleoanthropologist Christopher Stringer and others, which argues that H. sapiens arose in either eastern or southern Africa within the past 200,000 years and, because of its inherent superiority, subsequently replaced archaic hominin species around the globe without interbreeding with them to any significant degree. On the other was the Mulitregional Evolution model, formulated by paleoanthropologists Milford Wolpoff, Xinzhi Wu and Alan Thorne, which holds that modern H. sapiens evolved from Neandertals and other archaic human populations throughout the Old World, which were connected through migration and mating. In this view, H. sapiens has far deeper roots, reaching back nearly two million years. By the early 2000s the Recent African Origin model had a wealth of evidence in its favor. Analyses of the DNA of living people indicated that our species originated no more than 200,000 years ago. The earliest known fossils attributed to our species came from two sites in Ethiopia, Omo and Herto, dated to around 195,000 and 160,000 years ago, respectively. And sequences of mitochondrial DNA (the tiny loop of genetic material found in the cell power plants, which is different from the DNA contained in the cells nucleus) recovered from Neandertal fossils were distinct from the mitochondrial DNA of people todayexactly from the mitochondrial DNA of peopleexactly as one would expect if H. sapiens replaced archaic human species without mating with them. Not all of the evidence fit with this tidy story, however. Many archaeologists think that the start of a cultural phase known as the Middle Stone Age (MSA) heralded the emergence of people who were beginning to think like us. Prior to this technological shift, archaic human species throughout the Old World made pretty much the same kinds of stone tools fashioned in the so-called Acheulean style. Acheulean technology centered on the production of hefty hand axes that were made by taking a chunk of stone and chipping away at it until it made the desired shape. With the onset of the MSA, our ancestors adopted a new approach to toolmaking, nverting the knapping process to focus on the small, sharp flakes they detached from the corea more efficient use of raw material that required sophisticated planning. And they began attaching these sharp flakes to handles to create spears and other projectile weapons. Moreover, some people who made MSA tools also made items associated with symbolic behavior, including shell beads for jewelry and pigment for painting. A reliance on symbolic behavior, including language, is thought to be one of the hallmarks of the modern mind. The problem was that the earliest dates for the MSA were more than 250,000 years agofar older than those of the earliest H. sapiens fossils at less than 200,000 years ago. Did another human species invent the MSA, or did H. sapiens actually evolve far earlier than the fossils seem to indicate? In 2010 another wrinkle emerged. Geneticists announced that they had recovered nuclear DNA from Neandertal fossils and sequenced it. Nuclear DNA makes up the bulk of our genetic material. Comparison of the Neandertal nuclear DNA with that of living people revealed that non-African people today carry DNA from Neandertals, showing that H. sapiens and Neandertals did interbreed after all, at least on occasion. Subsequent ancient genome studies confirmed that Neandertals contributed to the modern human gene pool, as did other archaic humans. Further, contrary to the notion that H. sapiens originated within the past 200,000 years, the ancient DNA suggested that Neandertals and H. sapiens diverged from their common ancestor considerably earlier than that, perhaps upward of half a million years ago. If so, H. sapiens might have originated more than twice as long ago as the fossil record indicated.
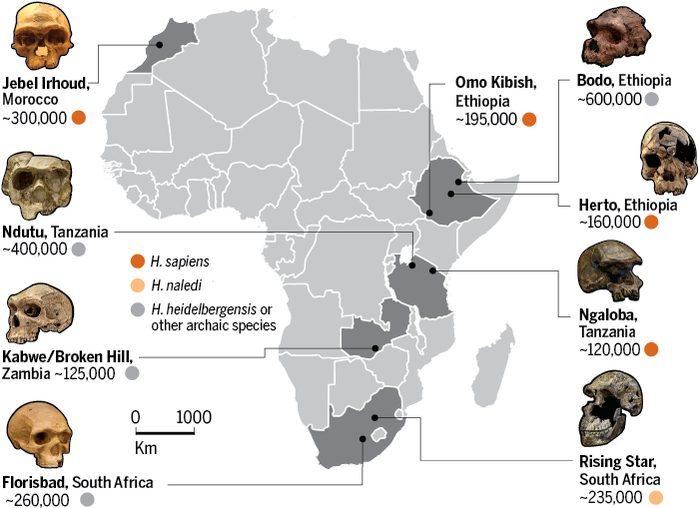
ANCIENT ROOTS Recent discoveries at a site called Jebel Irhoud in Morocco have helped bring the fossil, cultural, and genetic evidence into better alignmentand bolstered a new view of our origins. When barite miners first discovered fossils at the site back in 1961, anthropologists thought the bones were around 40,000 eyars old and belonged to Neandertals. But over the years continued excavations and analyses led researchers to revise that assessment. In June 2017 paleoanthropologist Jean-Jacque Hublin of the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany, and his colleagues announced that they had recovered additional fossils form the site, along with MSA tools. Using two dating techniques, they estimated the remains to be roughly 315,000 years old. The researchers had found the oldest traces of H. sapiens ot date, as well as the oldest traces of MSA culturepushing back the fossil evidence of our species by more than 100,000 years and linking it to the earliest known appearance fo the MSA. Not everyone agrees that the Jebel Irhoud fossils belong to H. sapiens. Some experts think they may instead come from a close relative. But if Hublin and his collaborators are right about the identity of the bones, the constellation of skull straits that distinguish H. sapiens from other human species did not all emerge in lockstep at the inception of our kind, as supporters of the Recent African Origin theory had supposed. The fossils resemble modern humans in having a small face, for example. But the braincase is elongated like those of archaic human species rather than rounded like our own dome. This shape difference reflects difference in brain organization: compared with fully modern humans, the Jebel Irhoud individuals had smaller parietal lobes, which process sensory input, and a smaller cerebellum, which is involved in language and social cognition, among other functions. Neither do the archaeological remains at Jebel Irhoud exhibit the full complement of MSA features. The people there made MSA stone tools for hunting and butchering gazelles that roamed the grasslands that once carpeted this now desert landscape. And they built fires, probably to cook their food and warm themselves against the chill of night. But they did not leave behind any traces of symbolic expression. IN fact, on the whole, they are not especially more sophisticated than the Neandertals fo H. heidelbergensis. If you could journey back in time to our species debut, you wouldnt necessarily pick it to win the evolutionary sweepstakes. Although early H. sapiens had some innovations, there werent any big changes at 300,000 years ago that indicate they were destined to be successful, observes archaeologists Michael Petraglia of the Max Planck Institute for the Science of Human History in Jena, Germany. In the beginning with sapiens, Petraglia says, it looks like anyones game.
GARDENS OF EDEN The total H. Sapiens package, many researchers agree, did not coalesce until sometime between 100,000 and 40,000 years ago. So what happened in the intervening 200,000 years or more to transform our species from run-of-the-mill hominin to world-conquering force of nature? Scientists are increasingly thinking about how the size and structure of the early H. sapiens population might have factored into the metamorphosis. In a paper published online in 2018 in Trends in Ecology & Evolution, archaeologist Eleanor Scerri of the University of Oxford and a large interdisciplinary group of co-authors, including Stringer, make the case for what they call the African Multiregionalism model of H. sapiens evolution. The scientists note that the earliest putative members of our speciesnamely, the Jebel Irhoud fossils from Morocco, the Herto and Omo Kibish fossils from Ethiopia, and a partial skull from Florisbad, South Africaall look far more different from one another than people today do. So they belong to different species or subspecies. But maybe early H. sapiens was just ridiculously diverse, Scerri offers. And maybe looking for a single point of origin for our species, as many researchers have been doing is a wild goose chase, she says. When Scerri and her colleagues examined the latest data from fossils, DNA and archaeology, the emergence of H. sapiens began to look less like a single origin story and more like a pan-African phenomenon. Rather than evolving as a small population in a particular region of Africa, they propose, our species emerged from a large population that was subdivided into different groups distributed across the vast African continent that were often semi-isolated for thousands of years at a time by distance and by ecological barriers such as deserts. Those bouts of solitude allowed each group to develop its own biological and technological adaptations to its own niche, be it an arid woodland, or a savanna grassland, a tropical rain forest, or a maritime coast. Every so often, however, the groups came into contact with one another, allowing for both genetic and cultural exchange that fed the evolution of our lineage. Shifting climate could have fueled the fracturing and rejoining of the subpopulations. For instance, paleoenvironmental data have shown that every 100,000 years or so, Africa enters into a humid phase that transforms the Sahara Desert into a lush expanse of vegetation and lakes. These Green Sahara episodes, as they are known, would have allowed populations formerly isolated by the harsh desert to link up. When the Sahara dried out again, populations would be sequestered anew and able to undergo their own evolutionary experiments for another stretch of time until the next greening. A population subdivided into groups that each adapted to their own ecological niche, even as occasional migration between groups kept them connected, would explain not only the mosaic evolution of H. sapiens distinctive anatomy but also the patchwork pattern of the MSA, Scerri and her co-authors argue. Unlike Acheulean tools, which look mostly the same everywhere they turn up throughout the Old World, MSA tools exhibit considerable regional variation. Sites spanning the time between 130,000 and 60,000 years ago in North Africa, for example, certain tool types not found at sites in South Africa from the same interval, including stone implements bearing distinctive stems that may have served as attachment points for handles. Likewise, South African sites contain slender, leaf-shaped tools made of stone that was heated to improve its fracture mechanicsno such implements appear in the North African record. Complex technology and symbolism become more common over time across the continent, but each group acts its own way, tailoring its culture to its specific niche and customs. H sapiens was not the only hominin evolving bigger brains and sophisticated behaviors, however. Hublin notes that human fossils from China dating between 300,000 and 50,000 years ago, which he suspects belong to Denisovans, exhibit increased brain size. And Neandertals invented complex tools, as well as their own forms of symbolic expressions and social connectedness, over the course of their long reign. But such behaviors do not appear to have become as highly developed or as integral to their way of life as they eventually did in ours, observes archaeologist John Shea of Stony Brook University, who thinks that advanced language skills allowed H. sapiens to prevail. All these groups are evolving in the same direction, Hublin says. But our species crosses a threshold before the others in terms of cognitive ability, social complexity, and reproductive success, And when it doesaround 50,000 years ago, in Hublins estimationthe boiling milk escapes the saucepan. Forged and honed in Africa, H. sapiens could now enter virtually any environment on the earth and thrive. It was unstoppable.
CLOSE ENCOUNTERS Hundreds of thousands of years of splitting up from and reuniting with members of our own species might have given H. sapiens an edge over other members of the human family. But it was not the only factor in our rise to world domination. We may actually owe our extinct relatives a substantial debt of gratitude for their contributions to our success. The archaic human species that H. sapiens met as it migrated within Africa and beyond its borders were not merely competitorsthey were also mates. The proof lies in the DNA of people today: Neandertal DNA makes up some 2 percent of the genomes of Eurasians; Denisovan DNA composes up to 5 percent of the DNA of Melanesians. And a recent study by Arun Durvasula and Siriam Sankararaman, both at the University of California, Los Angeles, found that nearly 8 percent of the genetic ancestry of the West African Yoruba population traces back to an unknown archaic species. Other genetic evidence from contemporary populations suggests that H. sapiens also interbred with unknown extinct hominins in South and East Asia. Some of the DNA that H. sapiens picked up from archaic hominins may have helped our species adapt to the novel habitats it entered on its march across the globe. When geneticist Joshua Akey of Princeton University and his colleagues studied the Neandertal sequences in modern human populations, they found 15 that occur at high frequencies, a sign that they had beneficial consequences. These high-frequency sequences cluster into two groups. About half of them influence immunity. As modern humans dispersed into new environments, they were exposed to new pathogens and viruses, Akey says. Through interbreeding, they could have picked up adaptations from Neandertals that were better to fight off those new pathogens, he explains. The other half of the Neandertal sequences that Akeys team found at high frequency in modern human populations are related to skin, including genes that influence pigmentation levels. Researchers have previously theorized that H sapiens individuals from Africa, who presumably had darker skin to protect against harmful ultraviolet radiation from the sun, would have had to evolve lighter skin as they entered northern latitudes to get enough vitamin D, which the body acquires mainly through sun exposure. Skin genes from Neandertals may have aided our predecessors in doing exactly that. Neandertals are not the only archaic humans who gave us useful genes. For example, modern day Tibetans have the Denisovans to thank for a gene variant that helps them cope with the low-oxygen environment of the high-altitude Tibetan plateau. And contemporary African populations have inherited from an unknown archaic ancestor a variant of a gene that may fend off bad bacteria inside the mouth. Interbreeding with archaic humans who had millennia to evolve adaptations to local conditions may well have allowed invading H. sapiens to adjust to novel environments faster than if it had to wait for favorable mutations to crop up in its own gene pool. But its not all upside. Some of the genes we obtained from Neandertals are associated with depression and other diseases. Perhaps these genes were advantageous in the past and only began causing trouble in the context of modern ways of life. Of maybe, Akey suggests, the risk of developing these diseases was a tolerable price to pay for the benefits these genes conferred. Archaic humans may have contributed more than DNA to our species. Researchers have argued that contact between divergent human groups probably led to cultural exchange and may have even spurred innovation. For example, the arrival of H. sapiens in western Europe, where the Neandertals long resided, coincided with an uncharacteristic burst of technological and artistic creativity in both groups. Previously some experts suggested that Neandertals were simply aping the inventive newcomers. But maybe it was the interaction between the two groups that ignited the cultural explosion on both sides. In a sense, the fact that H. sapiens mixed with other human lineages should not come as a surprise. We know from many animals that hybridization has played an important role in evolution, observes biological anthropologist Rebecca Rogers Ackermann of the University of Cape Town in South Africa. In some cases, it can create populations, and even new species, that are better adapted to new of changing environments than their parents were because of novel traits or novel combinations of traits. Human ancestors show a similar pattern: the combination of different lineages resulted in the adaptable, variable species we are today. Homo Sapiens is the product of a complex interplay of lineages, Ackermann asserts, and it has flourished precisely because of the variation that arose from this interplay. Without it, she says, we simply wouldnt be as successful. How often such mingling occurred and the extent to which it helped drive evolution in H. sapiens and other hominins remain to be determined. But it may be that the particular environmental and demographic circumstances in which our species found itself in Africa and abroad led to more opportunities for genetic and cultural exchange with other groups than our fellow hominins experienced. We got luckyand are no less marvelous for it.
Djehuti Member # 6698
posted
It should be obvious by now to everyone why the IBD-Yoruba West African marker should NOT be labeled as "Sub-Saharan".
^ The biggest division between Africans is between Aboriginal Southern Africans (Khoisan) and all other Africans.
This is why even Sub-Saharan West Africans (IBD populations) are closer related to West Eurasians than they are to Khoisan. As Loosdrecht et al. illustrated.
Another interesting fact is that the Khoisan folk are themselves genetically divided into two main groups as illustrated in Schlebusch's cladogram and was noted years ago.
Two studies exploring the genetics of hunter-gatherers and pastoralists in sub-Saharan Africa, where humans are thought to have originated, reveal that even though the click-language peoples of southern Africa live in close proximity, they belong to two distinct genetic clusters.
To assess the degree of genetic difference, both teams looked at single nucleotide polymorphisms (SNPs) variations at individual nucleotides between different people in the DNA of the various populations. One study, led by Carina Schlebusch at Uppsala University in Sweden and published today in Science1, analysed SNPs from 220 individuals belonging to 11 different southern African populations. The other, led by Joseph Pickrell at Harvard Medical School in Boston, Massachusetts, and posted to the arXiv preprint server ahead of its publication in Nature Communications2, examined SNPs from 187 individuals belonging to 23 southern and eastern African populations.
By considering the similarities and differences among the SNPs in the various click-speaking peoples, and comparing them with patterns from other African populations, the teams were able to identify ancestral relationships. Both teams deduced that the southern African click-speaking populations (known generally as the Khoisan) actually belong to two genetically differentiated groups, one in the north and and one in the south of the Kalahari, which went their separate ways around 30,000 years ago. The discovery of this genetic divide is raising numerous questions about how it could have come about.
It makes you wonder if they became isolated from one another for cultural reasons, or if there was some sort of geographical isolation that led to the differentiation, says Sarah Tishkoff, a human-origins geneticist at the University of Pennsylvania in Philadelphia.
Evolutionary geneticist Rasmus Nielsen at the University of California, Berkeley, agrees with the idea of the environment being involved. Africa was very dry during the last glacial maximum, and this might have led to an extended period of smaller overall population sizes, more population subdivision and increased fragmentation, he says.
East meets south
Aside from identifying a key time of separation among southern African peoples, Pickrell and his colleagues also discovered an ancient genetic relationship between the south and east of the continent. They found that the Sandawe and the Hadza, east-African hunter-gatherers who speak a click language but are geographically isolated from the other Khoisan peoples, derive roughly a quarter of their ancestry from the southern African click-speakers. This, Pickrell argues, implies the existence of a link having once existed between these peoples.
The possibility of such a connection is important because tensions have been growing among anthropologists as fossil evidence unearthed by palaeontologists increasingly suggests that modern humans originated in east Africa whereas genetic studies have suggested a southern origin. The identification of a genetic link between south and east Africa may help to resolve this conundrum.
Southern selection
Schlebusch and her colleagues also noticed that some of the genes in the populations they were studying seem to have undergone changes due to natural selection since the two southern-African groups split 30,000 years ago. They included genes involved in muscle function and growth and, intriguingly, genes involved in immunity.
The reasearchers found that the more southerly of the two groups carry evidence for selected changes in two immune-related genes, whereas these genes in the more northerly group show no signs of selection.
This signal could be due to this population having early and extensive contact with European colonists and suffering epidemics such as smallpox, explains Schlebusch.
Tishkoff says that this again raises the question of whether these populations were once geographically widespread and experienced local adaptation to different environments.
And as I've shown before, the dental nonmetrics very highly reflects genetics.
BrandonP Member # 3735
posted
quote:But such behaviors do not appear to have become as highly developed or as integral to their way of life as they eventually did in ours, observes archaeologist John Shea of Stony Brook University, who thinks that advanced language skills allowed H. sapiens to prevail. All these groups are evolving in the same direction, Hublin says. But our species crosses a threshold before the others in terms of cognitive ability, social complexity, and reproductive success, And when it doesaround 50,000 years ago, in Hublins estimationthe boiling milk escapes the saucepan. Forged and honed in Africa, H. sapiens could now enter virtually any environment on the earth and thrive. It was unstoppable.
I think this argument that advanced language capabilities and their ramifications distinguished modern humans (as in Late Stone Age/Upper Paleolithic descendants) from other hominins makes sense. I will have to dig it up later when I get home, but Chris Ehret has pointed out that even supposed AMH living in Israel over 100 kya would have had a vocal tract different enough from modern humans to have limited their linguistic capabilities. I do think 50 kya is a bit too recent to have been the origin point for LSA/UP humans though.
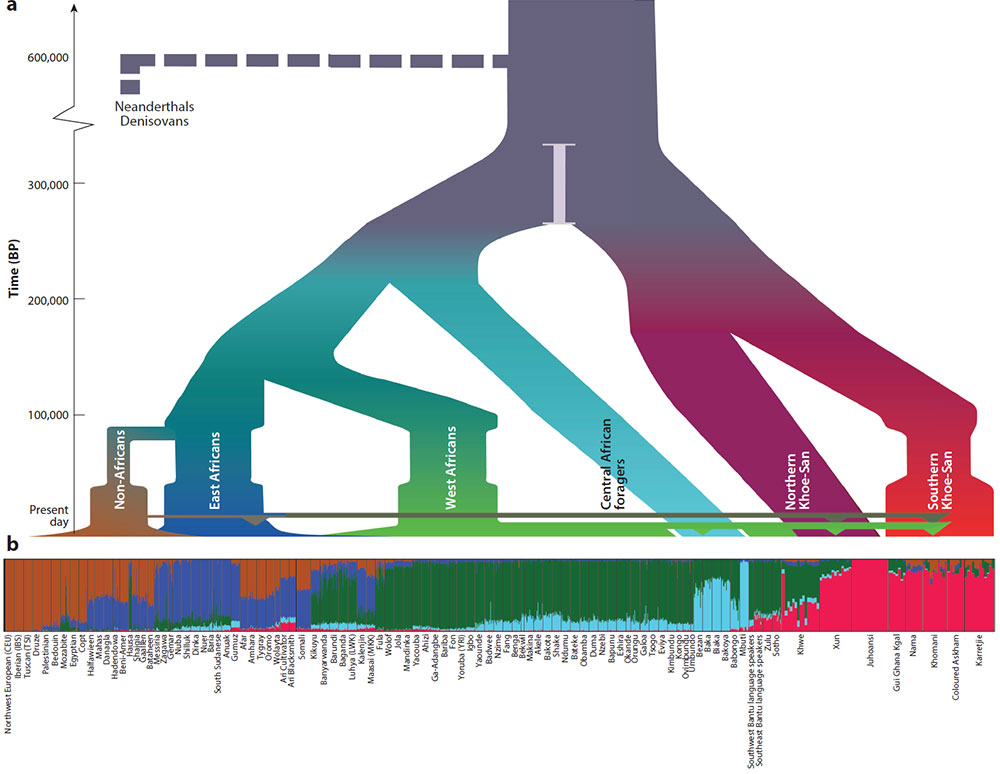
quote: Demographic model of African history and estimated divergences. (a) Population split times, hierarchy, and population sizes (summarized from 123). Horizontal width represents population size; horizontal colored lines represent migrations, with down-pointing triangles indicating admixture into another group. (b) Population structure analysis at 5 assumed ancestries (K=5) for 93 African and 6 non-African populations. Non-Africans (brown), East Africans (blue), West Africans (green), central African huntergatherers (light blue), and Khoe-San (red) populations are sorted according to their broad historical distributions. Data were obtained from several studies (4, 16, 44, 53, 70, 89, 125), and the details of the analysis are described in the Supplemental Methods . Abbreviation: BP, years before present.
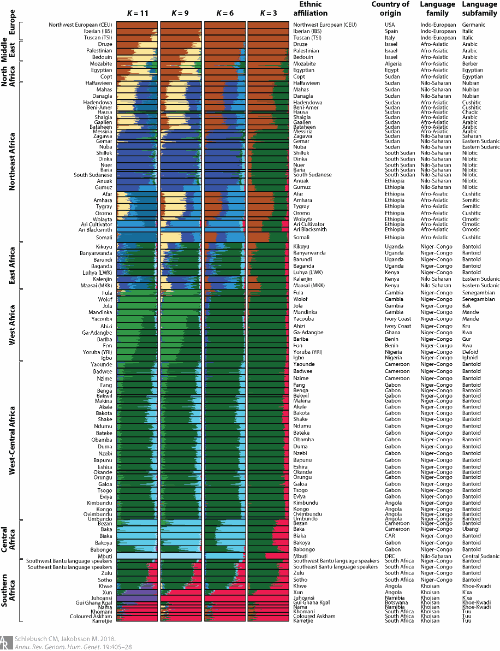
Figure 3
quote:
Population structure analysis and inferred ancestry components for selected choices of assumed number of ancestries (K=3, 6, 9, and 11) for 93 African and 6 non-African populations. Figure 2 displays the ancestry components for K=5. The broad geographical distributions are indicated on the left. The columns on the right indicate ethnic affiliation, country of origin, language family, and language subfamily. Data were obtained from several studies (4, 16, 44, 53, 70, 89, 125), and the details of the analysis are described in the Supplemental Methods . Abbreviations: CAR, Central African Republic; DRC, Democratic Republic of the Congo.
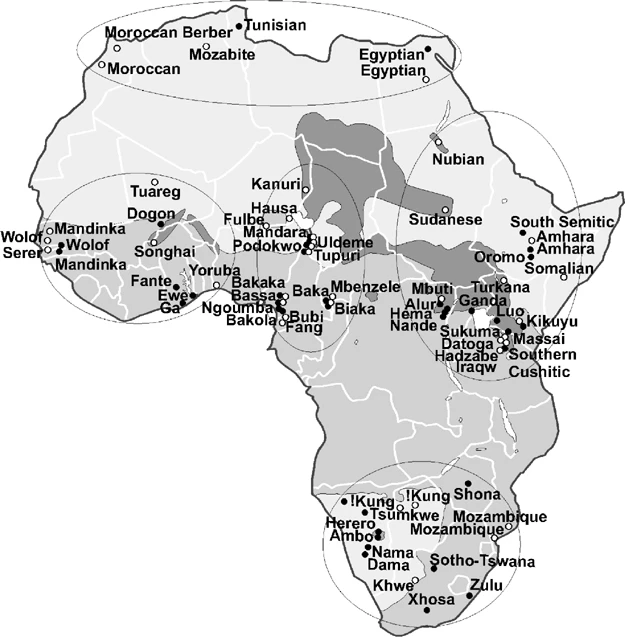
Interesting. I don't think i seen these before. Not to change the topic to much but i have to say however like i mention in another thread the mandinka term could be/is misleading because when folks think of mandinka at times they think of those of mali and guinea for example and that study above is really talking about the mandenka mandinka from gambia and not the maninka mandinka of mali and guinea.
When they use mandinka for the above i knew it was really gambia or senegal or both for these studies. Of course the above is talking only gambia.
The joshuaproject does not even list the maninka as mandinka in the People Name General/People Name in Country section. Note- but they do list mandinka has one of the alternate Names.
Alternate Names Konyanke; Mandingo; Mandinka; South Maninka; Wangara
This reminds me when folks get confused when nubian is talked about from sudan and egypt for example vs nubian from kenya because those nubians from kenya are not really nubians,they are nubi.
Then there is sudanese nuba vs nubian. Of course some nuba are really nubians(hill nubians) Then there is kushite vs cushite. One was a ethnic group and the other is a language group. Ancient ethiopia(sudan) vs ethiopia when the greeks were talking about ethiopia it was really kush/sudan.
posted
^ Thanks for the clarification on the Mandinka. I didn't know about the Senegambian Mandinka. I guess it's similar to how the agricultural Fulani of coastal Guinea are confused with the pastoral Fulani of the Sahel. I also had no clue about the Kenyan Nubians. This is why specificity is important in regards to population sampling.
Djehuti Member # 6698
posted
The paleoanthropologist Nutcracker Man has good articles.
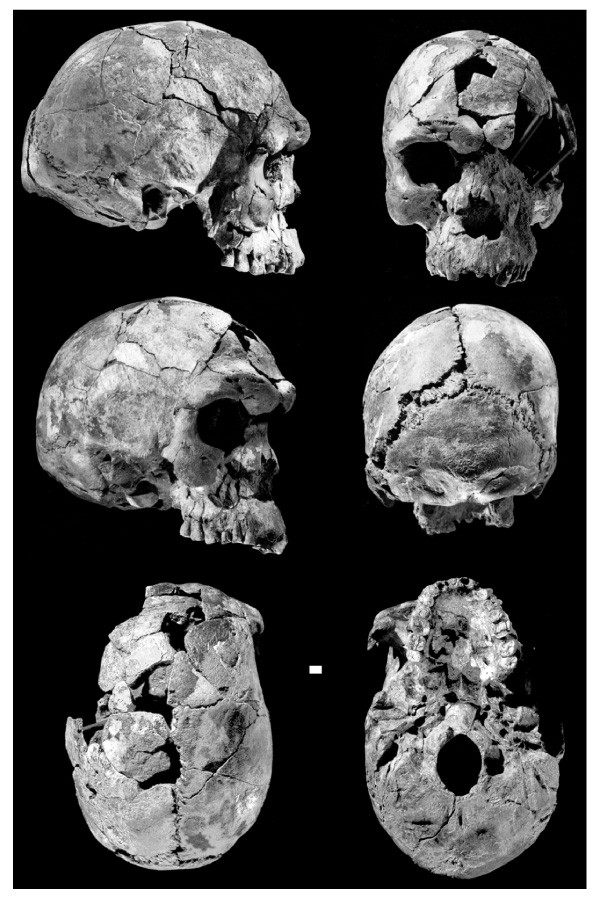
Many people forget that there was child found in the same site as Herto Man. Swenet and I discussed it here, but due to the relevance of this thread. I'll repost it below:
quote:Originally posted by Swenet: Must have been a rare report then, because there is almost no discussion on this child in the literature. Almost everything in the literature focuses on the adult Herto male. As a result the photographs of child's remains are far more difficult to find than the photographs of the adult male, even though both photographs come from the same paper. This is what it was like when I looked into this years ago. Situation might have improved in google, but I doubt it, because I know why it's like this in the first place.
I'm trying to find the paper I read years back when I was in high school, but I can only find recent papers like these:
African paleoanthropology has long been influenced by eurocentricframes of reference in geology, paleontology, and archaeology. Indeed, the Middle Stone Age of Africa is often equated with the Middle Pale-olithic as defined and manifested in Europe. Even the ill-defined Homo heidelbergensis is still applied to African fossils. More broadly, notions that technological cultures arose unilineally via transitions persist among Paleolithic archaeologists. On the other hand, and ironically, the strict application of cladistic classification disallows nomenclatural recognition of phyletic evolution along evolving species lineages. Combined with an under-appreciation of intraspecific skeletal variation, the approach has produced species inflation and a consequently confusing jungle of phylogenetic diagrams and labels applied to hominid fossils of all antiquities (White 2009, 2014). Scerri et al. represent a recent manifestation of this when they write of ...morphologically varied populations pertaining to the H. sapiens clade living throughout Africa(2018: 582). Left unspecified is what they mean by pertaining to. The inability of rigid biological classificatory labels to adequately express dynamic change continues to plague the depiction and under-standing of how our species lineage evolved through the Middle Pleistocene. However convenient static Linnean labels may be to claims about the earliest whatever taxon, such labels often work in opposition to expressing and understanding the mode and tempo of evolutionary change (White 2000, 2009, 2014).
For example, in biological anthropology a classificatory scheme based exclusively on branching speciation (cladistic classification, see below) would identify Homo sapiens based on an inferred Homo sapiens clade that reaches back >500,000 years ago, to a point at which African (Homo sapiens) and European (Homo neanderthalensis) biological species lineages diverged (apparently with minor subsequent introgression between them). In this scheme, anatomies as disparate as Bodo and Herto must both be classified cladistically as Homo sapiens.
Others adopt an anatomical approach, classifying as Homo sapiens only those fossils that display a specific suite of derived anatomical characters shared exclusively with anatomically modern humans. We prefer the latter approach, and nomenclaturally recognize those fossils (such as Bodo) interpreted to be the direct lineal ancestors of Homo sapiens as different chrono species, pending finer resolution of the paleontological record.
quote:Originally posted by Swenet: ^Did you notice that your paper has the Herto boy with recent humans, while the Herto adult is with so-called 'archaics'? That's exactly the evidence I wanted to see confirmed (see my second post where I said I'm waiting for clarification).
Yes! His juvenile morphometric pattern is similar to contemporary Sapiens juveniles. I was trying to find another study that came out right after the Herto discovery saying how because it's a child it is uncertain due to paedomorphic features showing similarity between different species of a genus or even family.
quote:Will have to go over it closely, to make sure I get everything from it. I also want to see if they elaborate on this massive morphological distance between these two Herto fossils and the fact that this means that adult Herto isn't in our lineage, at all. (imagine if Oase I is not ancestral to later Europeans/Eurasians, how far removed Herto must be given how similar he looks to the photographs of Kabwe in the 2003 White et al paper. [In the White et al paper the Herto adult, to me, looks like a cross/hybrid between the photographs of Qafzeh 9 and Kabwe, while Herto boy looks like neither and closer to us living humans]).
Yes, I have read that because of Herto's features he was deemed to probably come from a different line or subspecies of Sapiens hence his name Homo sapiens idaltu as opposed to Homo sapiens sapiens. However, I personally hesitate to make any conclusions without say conclusive genetic evidence. Recall the Zhoukoudian Upper Cave Series. All three skulls are contemporary to each other yet all three look very different from each other as to suggest different population origins yet the late Dr. Christy Turner and others have shown that all three skulls possess sinodont teeth. If there was that much craniometric variation among early East Asians at that time there is no telling what kind of variation existed in Africa that far back. If according to Turner the UC Zhoukoudian skulls belong to the same population despite their difference in facial features, who's to say that Herto Man and Child don't belong to the same family??!
quote:Originally posted by Swenet:
quote:Originally posted by Djehuti: Yes, I have read that because of Herto's features he was deemed to probably come from a different line or subspecies of Sapiens hence his name Homo sapiens idaltu as opposed to Homo sapiens sapiens. However, I personally hesitate to make any conclusions without say conclusive genetic evidence. Recall the Zhoukoudian Upper Cave Series. All three skulls are contemporary to each other yet all three look very different from each other as to suggest different population origins yet the late Dr. Christy Turner and others have shown that all three skulls are possess sinodont teeth. If there was that much craniometric variation among early East Asians at that time there is no telling what kind of variation existed in Africa that far back. If according to Turner the UC Zhoukoudian skulls belong to the same population despite their difference in facial features, who's to say that Herto Man and Child don't belong to the same family??!
Upper Palaeolithic sapiens skeletal remains at the same site often seem to belong to different 'races', either races we can recognize today (e.g. so-called 'Australoid' fossils), or they show variation large enough to be consistent with different races. You've mentioned the Upper Cave skeletal remains, and we also see it at Herto, Kostenki, Cro Magnon, Predmost, Qafzeh/Skhul, among other examples. Then there is the related finding that individual fossils separated by distance can have more of a resemblance than individuals at the same site. An example of this is Upper Cave 101 having its closest match in Ohalo I from the Levant, as opposed to anything at the Upper Cave site or the rest of China. Or you can have something like Mladec I showing more of a resemblance to Cro Magnon I than either does to Mladec II. Only at the latest phase of the Upper Palaeolithic we begin to see large samples that are homogeneous in a sense, in that they have a population affinity that is found more or less consistently throughout the population (e.g. Taforalt and Afalou, Jebel Sahaba, pre-Mesolithic al Khiday).
Scientists have never been able to explain this and I find it telling that some palaeontologists are going so far as to circulate new fossils amongst themselves before publishing so as to not get caught off guard by new fossils. You can't have it both ways. Either you're an expert and your predictions work and you're deserving of the trust the public vests in you, or you're not and you're losing control of the narrative with all these so-called archaics being found that don't match their predictions of human origins.
Firewall Member # 20331
posted
quote:Originally posted by Djehuti: ^ Thanks for the clarification on the Mandinka. I didn't know about the Senegambian Mandinka. I guess it's similar to how the agricultural Fulani of coastal Guinea are confused with the pastoral Fulani of the Sahel. I also had no clue about the Kenyan Nubians. This is why specificity is important in regards to population sampling.
Yes and here is some talk about that.
Gambian in Western Division Mandinka [GWD]
quote: Population Description
These cell lines and DNA samples were prepared from blood samples collected from people living in the Western District of Gambia from individuals who identified themselves as Mandinka. All parents in the trios identified themselves as having Mandinka parents of at least two generations.
Referring to Populations It is important to refer to this community as Gambian in Western Division Mandinka when describing these samples in articles or presentations. Including the full name reinforces the point that the sample set does not represent all Mandinka people or all Gambian people, whose population history is complex. The population should not be described merely as African, Sub-Saharan African, West African, or Gambian, since each of those designators encompasses many communities.
The full population descriptor is Gambian in Western Division Mandinka and the abbreviation is GWD.
Additional guidance about how to refer to the populations can be found at Guidelines for Referring to the Populations in Publications and Presentations.
Here are some population codes use in dna studies.
GF1 Fula I in Gambia
GF2 Fula II in Gambia
GMD Mandinka II in Gambia
GNA Akans in Ghana
GSR Serere in Gambia
GWD Gambian in Western Division, Mandinka
GWL Wollof in Gambia
MLB Bambara in Mali
MLM Malinke in Mali
MSL Mende in Sierra Leone
YRI Yoruba in Ibadan, Nigeria
quote: Map of Africa. The approximate location of 40 populations typed for Y chromosome markers in this study () and 39 populations surveyed for HVS1 sequence data12, 31, 32, 33 (○) are indicated. The distribution of the four African language families was constructed using Greenberg's39 classifications and further refined with data from the ethnologue (http://www.ethnologue.com/). Three shades of gray on map refer to the distribution of language families: Khoisan (light gray, southwest), Afroasiatic (light gray, north), Niger-Congo (medium gray), and Nilo-Saharan (dark gray). The circled geographic regions include North, West, Central, East, and South Africa.
quote:Originally posted by Djehuti: ^ If I understand you correctly Swenet, you consider the other AMH populations as what? Subspecies?? Anthropologists like Francis Howell, Tim White, and others, postulate that Herto Man due to his cranial features just being just slightly out of normal range of modern humans, to be instead to be part of its own subspecies-- H. sapiens idaltu as opposed to our subspecies H. sapiens sapiens.
In the thread you link to, you spoke of these other AMH lines possibly carrying mt lineage L3, yet there are modern West Africans who carry Y lineages that are much older-- A000.
Of course all these postulations are based on cranial traits alone. We need genetic evidence to be certain. We still don't know if the Herto child buried in the same grave and time period of Herto Man himself could be related despite the former showing very close affinities to modern sapiens sapiens children.
I personally wouldn't use a term like "subspecies" for a fossil that looks mixed, let alone raise a fossil to the level of 'subspecies' when its general morphology has not been attested anywhere else, and could very well be an isolated find that won't be repeated again.
Let's look at some of the things that have been said in the literature, and some of the things I keep in mind when I come to my conclusions.
1) According to Ehret, the Qahzeh-Skhul humans may not even have had the faculty of speech. So, if this is true, in my view, they're not nearly as close to us as 'experts' are claiming. And some of this likely extrapolates to other establishment-endorsed AMH.
In all early hominins, including the archaic humans in the Levant of 100,000 years ago, the pharynx extended downward at an oblique angle from the back of the tongue and was shorter than the oral cavity. In contrast, by 68 years of age in all fully modern human beings the pharynx descends at a right angle from the oral cavity, and the two segments of the vocal tract are of roughly equal length. All humans since 50,000 years ago possess this configuration.13 This modern human configuration has a high biological cost, however it makes fully modern human beings, unlike archaic humans, Neanderthals, and all other primates, susceptible to choking to death. But this configuration is also the essential enabler of our human ability to pronounce the full range of speech sounds, especially the primary vowels, which make modern human language possible. Clearly its benefits its enhancement of human communi- cation conferred survival benefits that outweighed its costs. 14 - Early humans: tools, language, and culture https://www.cambridge.org/core/books/abs/cambridge-world-history/early-humans-tools-language-and-culture/D607DC0C396C3A2807D93A72EB1D5E46
2) None of those establishment-endorsed AMH look morphologically modern in the way that Herto boy looks modern, even though Herto boy (160ky) is older than most of them. So, it seems to me that Herto boy, based on its age and morphology, should be used as a reference of what sapiens looked like ~160ky. How could it be possible to go from Omo (200ky) to essentially modern-looking Herto boy (160ky) in only 40ky? And then you have the fossils after that (e.g. Nazlet Khater), that do not look more modern than Herto boy. Based on this its crazy to try to organize this diversity of "anatomically modern" human forms (there are as many AMH forms in the Middle Palaeolithic as there as human fossils) into a lineage that leads to us, when you already have sth like Herto boy, which is already essentially like us, at this early 160ky date.
The childs cranium differs from the adult in being substantially smaller (especially with respect to maximum length) and has minimal development of its supra- orbital torus, and a much less projecting mastoid process and a smaller face, as one might expect in a juvenile. It also has very well-developed parietal bosses and a much more rounded contour of the external surface of the occipital. These details combine to endow BOU-VP-15/5 with an unequivocally modern morphology. Africa: The Cradle of Modern People https://catalogimages.wiley.com/images/db/pdf/9780470894095.excerpt.pdf
So, I'm not really worried so much about what those other 'AMH' are, as much as I'm confident in saying that they are not us (although, like Oase, who carries mtDNA N despite 10% Neanderthal introgression, they may still have varying degrees of sapiens ancestry). Those other fossils are disqualifying themselves by how 'archaic' they look compared to Herto and the Daoxin teeth. For instance, I already pointed out that, in the White et al 2003 paper, Herto adult looks like a mix between Kabwe and Qafzeh 9. Just like how O was able to link these Rift Valley skulls to different crania on the continent, you can learn a lot by doing the same thing with palaeolithic fossils.
3) According to multiregionalism exponents, there is a general lack of material evidence of an OOA migration 50-70ky ago.
3.1) You cannot find 'Cro-Magnons' in Africa 3.2) In most parts of the world, archaeological cultures show local continuation with earlier times and no sign of a universal, intrusive OOA migration bringing new technologies, and so on. 3.3) There is no clear Aurignacian/Upper Palaeolithic culture revolution in Africa, that can serve as a launching pad of OOA.
I don't agree with multiregionalism, but OOA exponents have never addressed these points. I myself can easily address these, based on my view that sapiens were a minority and are not to be identified with establishment-endorsed AMH that are dominant among the archaeological finds. Right away, this solves 3.1 and 3.3. But the establishment will never solve these because you cannot go from any of the AMH they endorse, to mtDNA N carriers in Europe who made Upper Palaeolithic tools. For instance, you cannot go from Aterians, to Upper Paleeolithic Europeans, nor can you go from LSA Africans, to Upper Palaeolithic Europeans. So, how do these 'experts' mean to solve these issues? They don't even acknowledge that multiregionalsts have a point that needs answering.
Djehuti Member # 6698
posted
^ I understand what you're saying now. Now that you mention it, I do recall the findings on the palate and pharynx shapes of many of these archaic AMHs not being adapted for speech the way those of modern sapiens-sapiens are which is the reason why I brought up the concept of sub-species and why I agree that all these archaic AMHs were NOT modern sapiens.
You also make some very salient points that I must concur with in regards to certain valid points that multiregionalists have been making all these years. It's as if modern Sapiens were not only a small minority but lurking in the background somewhere and all of a sudden just replaced all their cousins.
By the way, I first posted the paper here- The North African 'Negro' Paradox, but due to its relevance in this thread as well here it is again.
posted
Is it possible that LSA/UP humans absorbed all those so-called "archaic" populations elsewhere in Africa upon their expansion post-Toba? It seems likely to me that most of the latter were nonetheless closer genetically to LSA/UP than Neanderthals or Denisovans and therefore had more reproductive compatibility with us.
Djehuti Member # 6698
posted
^ That's what I'm thinking as well. I think the AMH introgression found in some Neanderthal samples actually come from these archaic AMHs rather than the more recent modern Sapiens, and that some of these Neanderthal admixed Archaics in turn mixed with modern Sapiens.
Swenet Member # 17303
posted
@DJ
quote:Originally posted by Djehuti: By the way, I first posted the paper here- The North African 'Negro' Paradox, but due to its relevance in this thread as well here it is again.
Interesting view (I didn't think of it that way, w/ negroid first in N. Africa, 'Caucasoid' first in Sub-Saharan Africa).
The way I see it, 'negroid', is not one thing, not even when you leave out 'negroid' Australoids, Veddoids, etc. and when you only focus on Africa. Now we have such a thing as ANA which can be described as SSA-like (Lawrence Angel's "Mouillian negroid"?), but not as Sub-Saharan African.
I would say about Vermeersch and Drobyshevsky that we don't really know what we're looking at when we see an African fossil like Nazlet Khater w/ negroid features. All we can say is that there is some type of distinctly African (ie non-Eurasian) ancestry in that fossil.
This unresolved part of African genetics doesn't just lead to conflation of differentiated populations (the danger of typological thinking), but it's also why Eurocentric bloggers are having a field day right now w/ Egyptian genomes.
BrandonP Member # 3735
posted
quote:Originally posted by Swenet: The way I see it, 'negroid', is not one thing, not even when you leave out 'negroid' Australoids, Veddoids, etc. and when you only focus on Africa. Now we have such a thing as ANA which can be described as SSA-like (Lawrence Angel's "Mouillian negroid"?), but not as Sub-Saharan African.
I would say about Vermeersch and Drobyshevsky that we don't really know what we're looking at when we see an African fossil like Nazlet Khater w/ negroid features. All we can say is that there is some type of distinctly African (ie non-Eurasian) ancestry in that fossil.
This unresolved part of African genetics doesn't just lead to conflation of differentiated populations (the danger of typological thinking), but it's also why Eurocentric bloggers are having a field day right now w/ Egyptian genomes.
Makes sense to me. If OOA and African substructure mean that some Africans will be genetically closer to Eurasians even without backmigrations, it would make sense for even sub-Saharan populations to vary with regards to Eurasian affinity, even if they all look "negroid" to a layperson.
Swenet Member # 17303
posted
What I mean is, we're now talking additional complexity that we now see play out even between neighbours (e.g. R12 Nubians and Gebel Ramlah Nubians, or seemingly Takarkori and Taforalt) where negroid morphology falls apart even more as a useful term, to the point where we can only use negroid features to say, there is distinctly African ancestry in a given population, but without being able to break it down further.
Ancient Nubians differentiating along hair texture lines, but not in terms of negroid features, could be further evidence of what I'm saying: that we don't know what ancestry underlies a skull with negroid features.
Djehuti Member # 6698
posted
quote:Originally posted by Swenet: What I mean is, we're now talking additional complexity that we now see play out even between neighbours (e.g. R12 Nubians and Gebel Ramlah Nubians, or seemingly Takarkori and Taforalt) where negroid morphology falls apart even more as a useful term, to the point where we can only use negroid features to say, there is distinctly African ancestry in a given population, but without being able to break it down further.
That's another reason why I believe so-called "Negroes" originate in North Africa, not only because the earliest type fossils occur there but such type fossils by the Holocene Era displays the greatest diversity traits. In fact anthropologists for decades have noted that 'Negroes' of the Sudan region (not the country) all across the Sahel have the greatest variation in contrast to Negroes further south.
quote:Ancient Nubians differentiating along hair texture lines, but not in terms of negroid features, could be further evidence of what I'm saying: that we don't know what ancestry underlies a skull with negroid features.
From what I understand Lower and Upper Nubians had the same cranio-facial form but the former had straighter hair while the latter had coiled hair.
quote:The way I see it, 'negroid', is not one thing, not even when you leave out 'negroid' Australoids, Veddoids, etc. and when you only focus on Africa. Now we have such a thing as ANA which can be described as SSA-like (Lawrence Angel's "Mouillian negroid"?), but not as Sub-Saharan African.
This is something I've suspected in regards to 'negroid' traits, but are you suggesting there could be other autosomal markers in North Africa (extant or extinct) that we don't know about??
quote:I would say about Vermeersch and Drobyshevsky that we don't really know what we're looking at when we see an African fossil like Nazlet Khater w/ negroid features. All we can say is that there is some type of distinctly African (ie non-Eurasian) ancestry in that fossil.
Note the PCA graphs above, particularly the one on ear bone structure. Hofmeyr is closer to Ishango than Nazlet Khater, though there is one EUP specimen that comes closest to Ishango.
quote:This unresolved part of African genetics doesn't just lead to conflation of differentiated populations (the danger of typological thinking), but it's also why Eurocentric bloggers are having a field day right now w/ Egyptian genomes.
This is exactly what I've been saying for months now. Say 'Basal Eurasian' and 'Natufian/Levantine'.
Elmaestro Member # 22566
posted
@Swenet I think what you're explaining is hard to grasp because the terminology to describe what we're looking at morphoogically etc. was already loaded. For instance Austrolasians AND contemporary black Africans being seen as "Negroid." So it's hard to make the connection if we look at some discrete traits which aren't categorically negroid (because of the populations who carried them more recently) that would only be seen almost exclusively in African populations going back in time. In this case, all modern Africans have mixed ancestry. We have dug up enough bones to have a good idea of who modern SSA's get their ancestry from, but it seems people are still holding out for a super-Negroid who would have mixed with Saharans, or something.
@Djehuti This is just about solved. I can say the latest neolithic Moroccan and fulani paper done allowed people to put 2 and 2 together, including me. What we consider SSA is a mixture which occured probably in the Central Sahara or Northern Congo for West/Central Africans and proto bantu (See Likasi individual Drennan 1942) and in North east African and the Sudan for what we consider Nilotics (See Becker 2012, pre-historic Wadi Howar paper)
Djehuti Member # 6698
posted
^ I provided Dr. Dobryshevsky's explanation of the 'Negroid complex' in my thread here.
Swenet Member # 17303
posted
@Elmaestro I agree. I try to tell ppl new to this. You might as well drop everything you know because it's not going to help you going forward. But luckily the stuff is not so difficult that you can't keep up. Discoveries (e.g. Takarkori's affinites) always have some ramifications down the line that can be used as clues/lessons (see Takarkori in graphic below). If you can catch those clues you can strike out on your own and put the literature/blogs on mute and still be correct.
@DJ Tried to be comprehensive, but all I had was paint, so excuse shitty graphics. You can start reading the Irish quote at the bottom. Then you can start reading the graphic below. Start with the balloon in the center from which the lines emanate and it should make sense.
This is what I mean with not meaningfully related:
MMD distances between the Neolithic and comparative samples are listed in Table 2. These inter-sample MMDs are also significantly different. It can be seen that Gebel Ramlah and R12 are phenet- ically less like each other than they are to the later comparative samples. A dental assessment of biological affinity between inhabitants of the Gebel Ramlah and R12 Neolithic sites.
Djehuti Member # 6698
posted
^ That graphic you made was exactly what I had in my mind, no lie! I had always thought it was more than coincidence that we find all these negroid features not just in North Africa but around the Mediterranean-- the Levant (Natufian), Neolithic Aegean, Malta, Iberia. Ironically, anthropologist Carleton Coon said such "negroid tendencies" were part of the 'proto-Mediterranean' stock.
Then we have the Ancestral North African autosomal marker which behaves like 'Sub-Saharan' in some ways but Eurasian in others.
^ They are a clear outlier compared to the other Nile Valley samples.
Just out of cursiosity, what do you make of the Garamantes?
Swenet Member # 17303
posted
@DJ If you remember, the initial Nea Nikomedea sample (n=4) had a nasal index pretty much the same as Mesolithic Nubians:
The Nea Nikomedeia from Greece [n=4] and Nubia 117 from southern Egypt manifest very similar nasal dimensions (59.4 and 59.7, respectively); however, while the high index in the Nubian group may be explained by Negroid gene flow into the population, the reason for such a high value in a European group is unknown. Artificial Skull Treatment in the PPNB Period: Nahal Hemar https://www.researchgate.net/publication/272719127_Artificial_Skull_Treatment_in_the_PPNB_Period_Nahal_Hemar
Yet this sample is Type B in affinities according to Briggs (ie no close affinities with Mesolithic Nubians):
Following the Palemediterraneans, the Af- rican Mediterraneans arrived, also from the Near East. In this case the physical type was a big and relatively rugged long-faced variant of the classic Mediterranean type, to which it is probably directly ancestral. We find people of this sort in Mesolithic sites in Palestine and in the lowest Neolithic levels of ancient Greece, in the Egyptian Pre-dynastic, at Elmenteita in Kenya and even farther south in Africa. Once again the African variety is dis- tinguished from its Near Eastern cousins by a series of minor and often seemingly negroid characters including relatively broad noses, sometimes with dull sills, some midfacial as well as alveolar prognathism, and, in a few cases, a peculiar and very distinctive man- dible of* the form that has been called "Boskopoid" (Briggs, 1955a, p. 63; pl. V, b). The Living Races of the Sahara Desert https://iiif.lib.harvard.edu/manifests/view/drs:53794813$1i
So you can see that the negroid contribution to the predynastics is different in that the nose is widened, but there is less corresponding shortening/widening of the face:
According to the indices, [Badarians] were dolichocranial, orthocranial, mesenic to leptenic and chamaerrhine (i.e. narrow, average height skull, average to narrow upper face, and rather broad nose). A rather high nasal index (men 54.8, women 55.2) together with marked prognathism distinguished them from the following Predynastic series and inclined towards the Negroid direction. Evidence of the Early Penetration of Negroes into Prehistoric Egypt https://www.jstor.org/stable/180563
And we can see that this checks out because Briggs' description of the small Nea Nikomedea sample, is supported by modern multivariate analyses, even though he came to his conclusions through visual observation, before the widespread adoption of statistical analysis by professionals in anthropology:
So, everything considered, there is a pristine Type B population (Olduvai-like?), and then there is one or more negroid populations that lived near and admixed with Type B in N. Africa (see that graphic I made). Based on the areas of the face especially affected by this admixture, we can get an idea of what this negroid ghost population looked like. For one, it must have been relatively long-faced compared to modern negroid populations.
And we can tell, as you pointed out, that northern Mediterranean samples from the Mesolithic (Muge) and Neolithic will have this admixture, even though talk of Natufian ancestry in Egyptians is all the rage now and no one is talking about the fact that predynastics and certain farmers were closer, morphologically (with only Shuqbah Natufians being closer, presumably).
quote:Just out of cursiosity, what do you make of the Garamantes?
Mainly Type B (though not the pristine Olduvai form which was presumably already assimilated by Iron Age times), but with a somewhat different cocktail of genetic components than Egyptians. BTW, that Wadi Ajjial sample in the dendrogram above, is from the Fezzan. So it has relevance to Garamantes.
quote:^ They are a clear outlier compared to the other Nile Valley samples.
I'm glad that older paper seems to support where I was taking my analysis with R12 Nubians. Sometimes you walk around w/ observations that are years old and really need revisiting/rechecking before you post them. Luckily my memory was on point.
Kanguet el Mouhaad 5 is Capsian and was used by Briggs as a good representative of the Type B female population. You can see how admixture has driven this individual's nasal index well into the Sub-Saharan African range and at the periphery of the Capsian samples (see the x axis), even though this individual does not have those affinities as much when you look at the total picture ( see Kanguet el Mouhaad 5). Like I said above, this tells us something about the populations involved in the admixture, and it shows how easy it is to get carried away by anthro reports talking about 'negroid' features and project modern Sub-Saharan Africans into those descriptions (which especially writers of African descent do) or to make them out to be ancestral to living Africans (which Vermeersch etc. appeared to be doing w/ Nazlet).
Djehuti Member # 6698
posted
^ I think the old Brauer study also does a good job of noting certain population affinities as well.
Brauer et al 1984
Swenet Member # 17303
posted
As far as Brauer, I got a lot out of his 1990 work with Rimbach, as well. See the 1st paper listed.
quote:Originally posted by Swenet: Have not read it yet. The only ones I use so far are from Brauer. I find most people don't understand them. Rightmire for instance thinks they are Bantu or Nilotes, as we discussed in a PM months ago. Some others think the that the phenotype originates from Eurasian ancestry brought to Kenya by pastoralists, even though Olduvai is 17ky and has no direct relationship to Kenyan pastoralists. Today most claim that they are a part of the Kenyan LSA, or Eburran, and that the Kenya Capsian lithic affinities were somehow mistaken/obsolete. But this has never been proved. But that's academia for you.
I find Brauer does a better job, although he uses terms like "Caucasian" and "Europid" to describe them, which is remarkable because he is one of the founders of OOA theory, so I feel he should know better as far as considering African substructure.
Brauer 1 (possibly the one Keita had in mind when he said "they cluster with Europeans") Late archaic and modern Homo sapiens from Europe, Africa, and Southwest Asia: Craniometric comparisons and phylogenetic implication https://www.sciencedirect.com/science/article/abs/pii/0047248490900213
The time period corresponds with the late part of the Nubian Complex (∼128,000 - 74,000 years ago).
Swenet Member # 17303
posted
@DJ Interesting. Is that the actual skull or is it a random stock photo?
If it's the actual skull, it looks much more 'modern' than the Taramsa Hill child burial from Egypt, which dates to around the same time.
quote:The skeleton appears to belong to an anatomically modern child. This is particularly evident from the morphology of the frontal bone which shows none of the recession or supraorbital development which would be expected in immature archaic humans at this developmental stage. Many features seem to he close to those of the robust Epipalaeolithic populations of North-Africa (Mechtoids) but also to those of the early anatomically modern humans of the Levant. The slenderness of the long bones, the rounding of the forehead and of the occipital region, the pentagonoid shape of the skull in occipital view and certain details of the orbits and their surroundings are undoubtedly anatomically modern features. On the other hand, the relatively large and apparently prognathic face may set this child closer to the more primitive forms of Jebel Irhoud, rather than the above-mentioned Mechtoid populations. Further comparisons with sub-Saharan Africa are necessary. On the basis of this preliminary assessment, an attempt to place this skeleton in a precise phylogenetic position would be dangerous. The fact that the skeleton belongs to a child who had not yet developed all the characteristics of an adult individual invites caution. Further study will be carried out on the surviving fragments after conservation and preparation.
Notice they say that the Taramsa Hill skull has a relatively large face, which, in this case, means, the face is relatively large compared to the forehead and brain case. This is the reverse of what we look for in sapiens. One morphotype of sapiens called the Boskop, which was found in South Africa, and, to a lesser extent, a somewhat related morphotype known as Cro-Magnon, are on the other end (large braincase, relatively smaller face). You can google 'Fishhoek skull' from South Africa, to get a good visual example of what that 'modern' combination of features looks like, when it's as extreme as in those fossils I've just mentioned.
Note Fishhoek on the right. Face is smaller, while in the 'archaics', the face is larger, and the large face is also combined with relatively smaller frontal bones (foreheads).
I'm no expert on all the details of how maturation into adulthood changes the expressions of archaic ancestry, but I do know that 'archaic' features still show up in 'archaic' juveniles. So, if that picture you posted shows a skull that belongs to Mtoto (ie if it's not a stock photo), you can see that, using the above pointers of 'craniofacial dysharmonie', as the French call it, the Mtoto skull seems more modern than Taramsa Hill from Egypt.
Though I'd like to have such observations on Mtoto seconded by someone knowledgeable who has actually studied the skull, before coming to conclusions. (Tut and KV55 turning out to be brachycranic, after decades of people calling attention to their 'dolichocranic Amarna headshape', shows how easy it is to get carried away w/ photographs).
Interestingly (and this is something that modern scholars are in denial about), this opposite condition to having archaic facial proportions with respect to the braincase, peaked in the palaeolithic, and it peaked especially in one subgroup of sapiens, but it doesn't peak in living populations anymore. Yes, believe it or not, the people who were the most modern in this respect, actually lived in the palaeolithic, and establishment-endorsed AMH often fall short of this important trait, that the establishment itself claims to use to define morphological modernity. Lol.
So, you can see why I don't go along with their eagerness to admit any halfway modern-looking fossil into 'AMH' or 'sapiens', as these fossils are really all over the map as far as morphological affinities, and it can't possibly be the case that all these fossils have a 200ky TMRCA in Omo I or a 300ky TMRCO in Irhoud, etc.
You might be interested in having another look at the graphic I posted earlier.
The y axis measures the craniofacial proportions I've mentioned above ('craniofacial dysharmonie'). You can see that Cro-Magnon I and Grotte des Enfants 4, a tall Gravettian known to belong to the Cro-Magnon morphotype, peak in this feature. Fishhoek and Boskop would also group in that bottom end of the spectrum, but numerous AMH and living humans would not (or not as much). You can also confirm that it checks out because, in the 1990 Brauer paper I mentioned above, Fishhoek clusters with Cro Magnon I, in one of the analyses (don't remember which PCA, but I'm confident it's in there).
Here is an older post I did on this:
quote:Originally posted by Swenet: As you know, there was a major recent announcement of convergence between certain UP remains in Africa and Eurasia. The convergence had a lot to do with rectangular orbits and what the French call 'craniofacial dysharmonie' (broadish and shortish face, coupled, ironically, with long and narrow neurocrania). From what I know, modern populations do not score high in this combination of traits. This is also shown in the paper (fig 4), where the UP remains stand out (concentrated in the upper left hand corner) due to their tendency to score high in these traits.
Hofmeyr and the UP Eurasian specimens tend to have comparatively high loadings on factor 2, which is indicative of a trend toward relatively longer crania with relatively shorter orbits than those in recent populations from these same geographic areas. Late Pleistocene Human Skull from Hofmeyr, South Africa, and Modern Human Origins
I now see this combination of traits, and certain others, as best describing the modal phenotype of the true (pristine) mtDNA N OOA population.
Update: Note that Herto child, already discussed above, also has the same relatively smaller face, unlike Herto adult:
The childs cranium differs from the adult in being substantially smaller (especially with respect to maximum length) and has minimal development of its supra- orbital torus, and a much less projecting mastoid process and a smaller face, as one might expect in a juvenile. It also has very well-developed parietal bosses and a much more rounded contour of the external surface of the occipital. These details combine to endow BOU-VP-15/5 with an unequivocally modern morphology. Africa: The Cradle of Modern People https://catalogimages.wiley.com/images/db/pdf/9780470894095.excerpt.pdf
Djehuti Member # 6698
posted
^ The photo I posted is a 3D model reconstruction based on computer scans of the actual remains as shown below.
According to the forensics the cranial proportions of the reconstruction have a high degree of accuracy, so I agree the Mtoto child does indeed look more modern than Taramsa, and is the oldest known Sapiens remains found in the Rift Valley.
Speaking of craniofacial development and paedomorphy, are you aware of the paedomorphic theory of human evolution?
quote:Neanderthal genes seen in modern humans may have entered our DNA through an interval of interbreeding starting about 47,000 years ago that lasted nearly 7,000 years, new research finds.
While there do seem to have been earlier dispersals (probably more than one) of AMH-related populations out of Africa, this new study suggests that the one modern OOA populations descend from happened not much earlier than 47 kya (as suggested by the date of admixture with Neanderthals).
quote:Stringer noted that prior research suggested that the interbreeding that introduced Neanderthal DNA into the modern human genome took place between 50,000 and 60,000 years ago. The new estimate of 47,000 years ago "has implications for Homo sapiens dispersals outside of Africa, because all extant [living] populations outside of Africa Chinese, Native Americans, Indonesians, Native Australians and so on carry the signs of this event, which therefore constrains when their ancestors began to disperse, to less than roughly 47,000 years ago," Stringer said.
However, "there is archaeological evidence of human occupation in northern Australia about 65,000 years ago," Stringer said. "So either that evidence is wrong; the populations were Homo sapiens but they went extinct or were swamped by a later dispersal; or the population was not, in fact, Homo sapiens." The latter possibility "seems much less likely given the complex behavior implied by the evidence, but would be a huge bombshell, of course."
Djehuti Member # 6698
posted
^ Swenet and I have discussed this a while back in regards to Pre-Toba vs. Post-Toba OOA expansions. It's probable the earlier Pre-Toba AM Humans were from an earlier line (subspecies) of Sapiens but not our own Sapiens sapiens lineage. Then again, it could be possible these were S sapiens but a branch that avoided admixing with Neanderthals. We see this in 'Basal Eurasian'. So unless we get some good OOA skeletal remains out of Africa it's anybody's guess.
Swenet Member # 17303
posted
@Brandon
While I respect Stringer's work and have appreciated some of his snaps comparing palaeolithic fossils on Twitter, the estimates he's talking about (assuming he's talking about LD estimates), are not as comprehensive as they seem.
Yes, the reported admixture dates from such methods seem reliable, but they don't give a full history of admixture events that we can detect in the uniparentals and in the ancient DNA. For instance, Armenian genomes report an admixture with a population related to Sub-Saharan Africans, that coincides with the arrival of 5.9 kiloyear bond event migrants ('Semitic speakers') from predynastic Egypt.
Admixture in the Near East
To formally test for population mixture in Armenians, we performed a 3-population test25 in the form of f3(Armenian; A, B), where a significantly negative value of the f3 statistic implies that Armenians descend from a mixture of the populations represented by A and B, chosen from the 78 global populations. We found signals of mixture from several African and Eurasian populations (Table 1, Figure 3). The most significantly negative f3 statistics are from a mixture of populations related to Sardinians and Central Asians, followed by several mixtures of populations from the Caucasus, Arabian Peninsula, the Levant, Europe, and Africa. We sought to date these mixture of events using exponential decay of admixture-induced LD. The oldest mixture events appear to be between populations related to sub-Saharan Africans and West Europeans occurring ~3800 bce, followed closely by a mixture of Sardinian and Caucasus-related populations. Later, several mixture events occurred from 3000 to 1200 bce involving diverse Eurasian populations (Table 1, Figure 3). Genetic evidence for an origin of the Armenians from Bronze Age mixing of multiple populations https://www.nature.com/articles/ejhg2015206
So, in short, what you do get is reliable, but you're not getting a full picture of the admixture events over time.
See how this method has performed in Egypt and in Morocco (see table 1 in Pagani 2012). Then compare these dates with the Naqada and Taforalt aDNA. The LD method detected a medieval admixture event with populations related to Sub-Saharan Africans. But, in reality, we know there has been sth SSA and SSA-like in Egypt and Morocco for much longer than the Middle Ages.
Taking this back to Stringer's comments. All we can say from the Neanderthal admixture event is that it happened. But we cannot say from this that older admixture events did not happen, or that older admixture events are not preserved in the genomes of modern Eurasians.
Swenet Member # 17303
posted
quote:Originally posted by Djehuti: ^ The photo I posted is a 3D model reconstruction based on computer scans of the actual remains as shown below.
Too bad reconstructions of that type (ie from scratch--literally) would probably not be considered admissible evidence. Would have been a great addition to this case I'm building around Herto boy, which I think would really benefit from securely dated and well preserved remains.
quote:Originally posted by Djehuti: ^ Swenet and I have discussed this a while back in regards to Pre-Toba vs. Post-Toba OOA expansions. It's probable the earlier Pre-Toba AM Humans were from an earlier line (subspecies) of Sapiens but not our own Sapiens sapiens lineage. Then again, it could be possible these were S sapiens but a branch that avoided admixing with Neanderthals. We see this in 'Basal Eurasian'. So unless we get some good OOA skeletal remains out of Africa it's anybody's guess.
Personally, I do not know which non-sapiens human population is truly a subspecies and which is not. Sapiens do seem to be a subspecies in their own right, since they cannot be easily conceived as arising out of an admixture event with any so-called 'archaics' known to us. (Sapiens brain size, and sapiens modern behaviour alone, would seem to rule that out).
But Neanderthals can definitely be conceived as arising out of an admixture event involving sapiens and Sima de los Huesos. So, I would not assign Neanderthals to a subspecies, since they are not a subspecies in their own right (not to me, at least, but others can have their own view on that), and most AMH I also would not refer to as a subspecies (since they seem to be to be a product of admixture, rather than emerging as a population, independent of genetic input from sapiens).
For instance, it would take a lot for me to accept that the mixed pattern of 'archaic' and more 'modern' features of homo naledi did not result from admixture. So, if it's the first step to more presumptuous speculation (e.g. elevating them to a subspecies in their own right, or putting them on their own branch in the human tree), I would not accept from the literature the term 'homo naledi' as anything other than a man-made construct, invented by someone with a phd, sitting behind a desk. But, like I said, others are free to make of such actions by the literature (ie the unchecked proliferation of species and subspecies constructs in the absence of DNA), what they want to.
So, when I talk about these extinct palaeolithic populations that the scientific literature calls 'AMH' or 'archaic' or 'transitional', I try to only use words like 'sapiens', for us, and hybrid or non-sapiens for fossils that seem only partly related to us, through admixture.
Djehuti Member # 6698
posted
^ I get what you're saying. There are experts who think Herto Man to be a subspecies known as Homo sapiens idaltu while our own kind is H. sapiens sapiens. I remember in one of my biology classes in university there was a discussion on what exactly is the line between subspecies and 'breed'-- that is population isolate that is still able to interbreed with similar populations. You see this in multiple species. As an example with canines, it was thought for a long time that domestic dogs descended directly from gray wolves due to the close genetic resemblance and ability to interbreed. The latest genetic analysis however instead shows that modern dogs do not descend directly from gray wolves but rather from a distinct line that happens to share recent common ancestry with them. Similarly red wolves show the same type of relation with coyotes-- all are in the genus Canis. I'm wondering if the same is true with Homo with some species being more related to others. So perhaps Herto Man can be thought of as a species that is genetically close (?)
Swenet Member # 17303
posted
I didn't know that about dogs, but if that's true, that would be just another example of scientists making elaborate phylogenetic trees, without any DNA.
BTW, it's funny you bring this up, because weeks ago I was looking into the historical differences between amstaffs and pitbulls, and I was surprised at how trivial the differences are between the two breeds, especially considering how how people are often defending these differences, as being hard boundaries. But the scale of differences they're debating, we're not used to, on here. The scale of difference they're talking is, like, Otzi (EEF) and Stuttgart (EEF), or maybe even a smaller difference that that. The way I see it, differences between the dogs I mentioned, is nothing like the genuine physical differences between, say, a poodle and a rottweiler. In my opinion, this just shows you how people professing to talk biological realities, can get carried away and really just push nonsense.
So, yes, I agree with what you said above. But in my discussion of 'subspecies', the direction I was taking it, was slightly different. What I meant, is this. Consider these statements often made by scholars. I'm sure you recognize at least some of them:
"Neanderthals, at times, made tools just as complex as the tools made by sapiens" "Neanderthals had a brain size right on par with sapiens, or they had an even larger brain size" "Neanderthals were capable of making flutes, which proves they made complex tools" "If we'd dress a Neanderthal up in a modern business suit, they wouldn't stand out if you passed them by, on the streets, today" "Neanderthals are our distant cousins and our closest relatives on the human tree; we separated from them, only ~400ky ago"
Statements like this betray an underlying premise that Neanderthals are a population in their own right, essentially independent from sapiens, after they branched off, 400ky ago. Otherwise, if they acknowledged that Neanderthals could have massive infusion of sapiens DNA, these types of conversations would be much more complex, and it would be acknowledged that merely finding flutes or advanced tools in Neanderthal Europe, is not evidence that Neanderthal made them. So, you can tell, the whole premise, the whole agenda is to treat these archaics as subspecies in their own right, in terms of being on a parallel 'trajectory' from sapiens.
My point is, I don't buy any of this in relation to Neanderthals (and I'm considering that the same could also be bogus for some other so-called 'archaics' [I just don't have the aDNA of erectus, etc. to prove it]). We know now, that Neanderthal uniparental profiles are filled with AMH haplogroups, and that they've lost their local European or Denisovan-like haplogroups. So, we know that the above premise is just a reflection of phds sitting behind their desks, dreaming up constructs and coming up with narratives that are not rooted in evidence.
This is the underlying point I was trying to make about rejecting scientists giving out species and subspecies designations, like Oprah giving out gifts to members in her audience. This is also why I don't buy the "home sapiens idaltu" designation. We can clearly see in that White et al paper, that Herto adult looks mixed between something Kabwe, and something Qafzeh 9.
Djehuti Member # 6698
posted
^ Indeed. I actually find it humorous how academia initially made the Neanderthals as dull-witted sub-humans who succumbed to extinction due to failing to compete with the superior Sapiens that came in, but when it came to light that many of these Sapiens show Neanderthal admixture and that these Neanderthals themselves show Sapiens haplogroups, then all of a sudden they were made into brilliant beings who would look no different from a Sapiens when dressed in modern clothes. The last part makes no sense considering the considerable differences in anatomical traits.
Swenet Member # 17303
posted
^I look forward to see how this develops.
quote:Originally posted by Swenet: "Neanderthals, at times, made tools just as complex as the tools made by sapiens"
^I also would like closure (ie secure dates) on the Chatelperronian, a tool industry once thought to date to Neanderthal (ie pre-Aurignacian) times, even though elements of the tools recall African features that would go against this forced interpretation of treating anything modern looking in Neanderthal Europe as Neanderthals being on an independent and entirely parallel track to modern humans, as far as speciation. I know these same scholars must be seeing that elements of the Chatelperronian seem African (the Chatelperronian has, in fact, been linked to a Mesolithic African incursion into Europe), but ignoring inconvenient evidence seems entrenched in the MO of mainstream academia.
Either way, if the Chatelperronian is indeed older than the mtDNA N-linked migration wave, this is just one more checkmark ✓ in my checklist of evidence that sapiens are mostly going unnoticed in the fossil record (that is, until their numbers grew with expansions of mtDNA L3, mtDNA N, and mtDNA M), and it's just more evidence that sapiens are not to be conflated with establishment-endorsed AMH, like Herto male and Omo (who never made anything comparable to the Chatelperronian).
I know I'm not the only one scratching his head at what is coming out of mainstream palaeontology sometimes (I know some lurkers have to be feeling me on this open bs by the establishment). But, like I said, I'm looking forward to see where this academic hypetrain, that everyone is buying into, is going.
BrandonP Member # 3735
posted
quote:Originally posted by Swenet: Either way, if the Chatelperronian is indeed older than the mtDNA N-linked migration wave, this is just one more checkmark ✓ in my checklist of evidence that sapiens are mostly going unnoticed in the fossil record (that is, until their numbers grew with expansions of mtDNA L3, mtDNA N, and mtDNA M), and it's just more evidence that sapiens are not to be conflated with establishment-endorsed AMH, like Herto male and Omo (who never made anything comparable to the Chatelperronian).
Unfortunately, the African tropics aren't an optimal place to preserve organic remains for extended periods of time due to the heat and humidity. Is it possible that the forerunners of LSA/UP humans were at one point concentrated in a part of Africa where the climate was not ideal for preserving our skeletal remains?
Swenet Member # 17303
posted
^I'm sure that plays a role, as well. But sapiens left behind their Y-DNAs and mtDNAs in the genepools of Neanderthals, which is one big breadcrumb showing us where they went. And we don't have much in the way of sapiens bones in cold Neanderthal Europe, either (so, the bones are not just lacking in warm equatorial Africa).
It seems to me that the world was just very hostile, and that things like the Toba eruption were need to kill off large swaths of archaics so that sapiens could increase there numbers more easily, after things recovered in the generations after the fallout from the eruption.
I believe this is why mtDNA L3, M and N blow up after the Toba eruption, ~70ky ago, and why humans were concentrated more in refugia, before that (like I said, I think it wasn't safe).
That's my take, for which I have more reasons than what I've just said.
One reason being, that Egypto-Nubians also couldn't take the Nile until very late, even though we know from Briggs observations of Iberomaurusians, from al Khiday, from Shuqbah, etc. that ancestors of Egypto-Nubians were already in the area. Just not at the Nile (not for long, anyway, for those that did venture there). The Maghreb was also taken late from Aterians and Aterian-like leftovers. So you can see patterns repeating in different areas formally held by humans of various kinds, who were not our ancestors, and usually you also see environmental catastrophes (that would take smarts to survive, therefore killing the non-sapiens grunts) precede these takeovers.
Djehuti Member # 6698
posted
^ I recall a thread started some time back about evidence of an early or earlier wave of OOA into Europe via the Straits of Gibraltar. I have always suspected of there being earlier waves of OOA prior to the 'main' one and that some of them entered directly into Europe either via the straits of Gibraltar OR by island hopping. Indeed the Mediterranean Basin was much shallower during the LGM, so that could explain things. However, I do find it strange that there is a large gap in the fossil record. But I agree that perhaps a major reason why Sapiens did not successfully colonize Eurasia pre-Toba was due to too much competition with other Hominids not to mention the megafauna that existed in that supercontinent. Funny how back in my zoology class in university, I learned about how many species of fauna especially megafauna became extinct during the Pleistocene due to environmental changes and/or introduction of new invasive species, with the latter being a likely consequence of the former. I never thought much about the situation being the same for Sapiens. As the OP article I cited as well as many other studies suggest then Africa truly was the cradle of Sapiens being a safe haven compared to the harsher Eurasian landscape.
By the way, in regards to fossils if I recall accurately the Aterians are to be identified with the Dar-es Soltan skull which is a true Sapiens and not the earlier Jebel Irhoud or Temara child which represent other lines or species??
Swenet Member # 17303
posted
Good point with the megafauna. Will refresh my memory on what that means in the Middle and Upper palaeolithic, other than the obvious (e.g. mammoths, and aurochs).
As far as the Maghreb, I consider both Irhoud and DS-5 as being not us, although the latter I'm sure has a significant infusion of sapiens ancestry.
See short-lived signs of sapiens presence in Maghreb, before assimilation with non-sapiens and the inevitable extinguishing of the full suit of modern behaviour (same thing you see at the Nile, where there is no art, until the Silsilian; then the art disappears again, as the Silsilian disappears).
So, this is not difficult. You can see why I avoid calling establishment-endorsed AMH, sapiens. Sapiens were there, in the Maghreb (worked bone, among other signs), but they were eventually assimilated, which you can see, also, in DS5 being closer in affinity to the modern samples, than Irhoud is (compare, for instance, Irhoud's relative closeness to archaics in the distance tables; although it's not easy to see in the dendrograms).
EDIT: Looks like, based on these facial variables, at least, Irhoud is closer to both archaics and moderns in the above Harvati and Hublin paper, than DS-5 is. Confusing result. Will check other papers later on.

 posted
posted