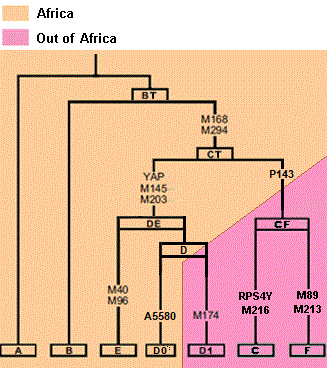
E-P2 (PN2) is one of the main haplogroup in Africa along with other E, A and B haplogroups.
It has its origin after the main OOA migration of non-Africans. E-P2 carriers weren't part of the main OOA migration.
So E-P2 unites African people and is probably one of the main haplogroup among Ancient Egyptians along with other A, B and E haplogroups.
Most Niger-Congo(Kordofanian), Cushitic and Chadic speakers are carriers of E-P2.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote: Using the principle of the phylogeographic parsimony, the resolution of the E1b1b trifurcation in favor of a common ancestor of E-M2 and E-M329 strongly supports the hypothesis that haplogroup E1b1 originated in eastern Africa, as previously suggested [10], and that chromosomes E-M2, so frequently observed in sub-Saharan Africa, trace their descent to a common ancestor present in eastern Africa.
-- from A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms (Trombetta 2011)
Y chromosome diversity, human expansion, drift, and cultural evolution Jacques Chiaroni,a Peter A. Underhill,b and Luca L. Cavalli-Sforzac,1 2009
Haplogroups A and B are the deepest branches in the phylogeny and are essentially restricted to Africa, bolstering the evidence that modern humans first arose there (14, 15). Haplogroup A is mainly found in the Rift Valley from Ethiopia to Cape Town, mostly but not exclusively in some of the oldest hunter-gatherers who still survive and speak Khoikhoi and San languages, proposed by some to be the oldest languages. The interruption of its distribution in the middle of the Rift Valley is possibly the consequence of replacement by Bantu-speaking farmers who settled the region starting in the first millennium of the Christian era. Haplogroup B is found mainly among African Pygmies, who live in the central African forest and are still predominantly hunters-gatherers but speak Bantu languages borrowed from farmers who arrived in the area between 2,000 and 3,000 years ago. The third predominantly African haplogroup, E, diversified some time afterward, probably descending from the East African population that generated the Out of Africa expansion. The geographic distributions of the major branches of this haplogroup, given in Fig. S1b, suggest that most of the settlement outside of Africa by haplogroup E members involves the later mutant E-M35 varieties like M78, M81, and M123 that extended to Arabia and the northern Mediterranean coast.....
It is also interesting to sum the distributions of different haplogroups descending from the same mutation, as for example D and E, which both descend from DE-YAP, the first mutation that split into the E branch that perhaps returned to Africa (or arose there), whereas the other branch, D, is found today mainly in the Himalayas and Japan.
Posted by Trollkillah # Ish Gebor (Member # 18264) on :
^for the trolling person above. It's funny and pathetic how you fight this with every drop of blood running through your racist veins.
Sub clade E-M81 is known as the Berber clade, since it's regional to Northwest Africa.
And for Yap DE*.
quote:The DE haplogroup appeared approximately 50,000 years bp in North East Africa and subsequently split into haplogroup E that spread to Europe and Africa and haplogroup D that rapidly spread along the coastline of India and Asia to North Asia.
quote: The BT haplogroup split from the root of the Y haplogroup tree 55,000 years before present (bp), probably in North East Africa. The CF(xDE) haplogroup was the common ancestor of all people who migrated outside of Africa until recent times. The defining mutation occurred 31-55,000 years bp in North East Africa and is still most common in Africa today in Ethiopia and Sudan. The DE haplogroup appeared approximately 50,000 years bp in North East Africa and subsequently split into haplogroup E that spread to Europe and Africa and haplogroup D that rapidly spread along the coastline of India and Asia to North Asia.
quote: "Firstly, haplogroup E-M2 (former E1b1a) and haplogroup E-M329 (former E1b1c) are now united by the mutations V38 and V100, reducing the number of E1b1 basal branches to two. The new topology of the tree has important implications concerning the origin of haplogroup E1b1. Secondly, within E1b1b1 (E-M35), two haplogroups (E-V68 and E-V257) show similar phylogenetic and geographic structure, pointing to a genetic bridge between southern European and northern African Y chromosomes. Thirdly, most of the E1b1b1* (E-M35*) paragroup chromosomes are now marked by defining mutations, thus increasing the discriminative power of the haplogroup for use in human evolution and forensics."
[...]
Within E-M35, there are striking parallels between two haplogroups, E-V68 and E-V257. Both contain a lineage which has been frequently observed in Africa (E-M78 and E-M81, respectively) [6], [8], [10], [13][16] and a group of undifferentiated chromosomes that are mostly found in southern Europe (Table S2). An expansion of E-M35 carriers, possibly from the Middle East as proposed by other Authors [14], and split into two branches separated by the geographic barrier of the Mediterranean Sea, would explain this geographic pattern. However, the absence of E-V68* and E-V257* in the Middle East (Table S2) makes a maritime spread between northern Africa and southern Europe a more plausible hypothesis. A detailed analysis of the Y chromosomal microsatellite variation associated with E-V68 and E-V257 could help in gaining a better understanding of the likely timing and place of origin of these two haplogroups.
--Beniamino Trombetta, Fulvio Cruciani et al. (2011)
A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms
There ya go!
Posted by the lioness, (Member # 17353) on :
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Trollkillah # Ish Gebor: [QB] ^for the trolling person above. It's funny and pathetic how you fight this with every drop of blood running through your racist veins.
Sub clade E-M81 is known as the Berber clade, since it's regional to Northwest Africa.
what I posted supports much of what Amun Ra posted so what are you talking about?
Diop, Clyde and Amun Ra are more Afrocentric that you are. You cliam that berbers are primarily African. They don't. And this is bourne out by the maternal DNA
Posted by xyyman (Member # 13597) on :
Nice synopsis TP.
@ Lioness. So...We are back to White women leaving their white men behind...20ya...in search of black men. Or were they hunting in Africa for food and decided to stay and not go back?
I know some brothas like to think they are all that but...
Posted by xyyman (Member # 13597) on :
Can someone explain this? I am not sure what they are trying to say here...
--- The IJ haplogroup characterizes part of the second wave of emigration from Africa that occurred via the Middle East 45,000 years bp and defines two branches I and J that emigrated northwards and eastwards into Europe. The J branch subsequently split again and contributed to the current North African population. The NO haplogroup appeared approximately 35,000-40,000 years bp in a region east of the Aral sea; subsequent branches spread to North Asia (N) and another branch (O) to South Asia via North India
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by xyyman: Nice synopsis TP.
@ Lioness. So...We are back to White women leaving their white men behind...20ya...in search of black men. Or were they hunting in Africa for food and decided to stay and not go back?
I know some brothas like to think they are all that but...
It's called rape and pillage
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by xyyman: Can someone explain this? I am not sure what they are trying to say here...
--- The IJ haplogroup characterizes part of the second wave of emigration from Africa that occurred via the Middle East 45,000 years bp and defines two branches I and J that emigrated northwards and eastwards into Europe. The J branch subsequently split again and contributed to the current North African population. The NO haplogroup appeared approximately 35,000-40,000 years bp in a region east of the Aral sea; subsequent branches spread to North Asia (N) and another branch (O) to South Asia via North India
In human genetics, Haplogroup IJ (S2, S22) is a human Y-chromosome DNA haplogroup. Haplogroup IJ is a descendant branch of the greater Haplogroup F (M89, P14, M213).
It is notable that no example of a Haplogroup IJ* Y-chromosome has been found among any modern human population; the existence of the Haplogroup IJ node has been inferred from the fact that certain mutations are shared in common among all Y-chromosomes belonging to the descendant haplogroups I and J. The lack of any examples of Haplogroup IJ* belonging to neither Haplogroup I nor Haplogroup J complicates any attempt to deduce the geographical location where Haplogroup IJ first appeared; however, the fact that both Haplogroup I and Haplogroup J are found among modern populations of the Caucasus, Anatolia, and Southwest Asia tends to support the hypothesis that Haplogroup IJ derived from Haplogroup F in the vicinity of West Asia or the Middle East and subsequently spread throughout Western Eurasia.
In human genetics, Haplogroup I (M170, P19, M258) is a Y-chromosome haplogroup. According to current theories, Haplogroup I first arrived in Europe around 20,000-25,000 years ago from the Middle East, perhaps associated with the Gravettian culture[2], and just prior to the onset of the last glacial maximum (LGM). It is most closely related to Haplogroup J,the "Semitic marker" being the most common in Semitic peoples, as both Haplogroup I and Haplogroup J are descendants of Haplogroup IJ (S2, S22). Haplogroup IJ is in turn derived from Haplogroup F. Haplogroup I (the letter I, not the number 1) can be found in most present-day European populations, most commonly in Scandinavia, Sardinia, and the Slavic &Bulgarian populations of the Western Balkans in southeastern Europe.
The distribution of J1 outside of the Middle East may be associated with Arabs and Phoenicians who traded and conquered in Sicily, southern Italy, Spain, Azerbaijan, Turkey, and Pakistan, or with Jews, who have historical origins in the Middle East and speak (or historically spoke) a Semitic language, though typically Haplogroup J2 is more than twice as common among Jews.
Haplogroup J2 It is composed of several sub-Haplogroups representing several different countries like Turkey, Iraq, Kurdistan, Lebanon, Syria, Armenia, Georgia, Aegean, Balkan, Italy. One sub Haplogroup M172* is mainly found in the Northern Fertile Crescent, the Mediterranean, Iran, Central Asia, and Southern Europe. Is is though to have originated in Anatolia (Turkey and Kurdstan) ie North Mesopotamia, and spread to Europe and to other Middle countries
Posted by Clyde Winters (Member # 10129) on :
quote:Originally posted by xyyman: Can someone explain this? I am not sure what they are trying to say here...
--- The IJ haplogroup characterizes part of the second wave of emigration from Africa that occurred via the Middle East 45,000 years bp and defines two branches I and J that emigrated northwards and eastwards into Europe. The J branch subsequently split again and contributed to the current North African population. The NO haplogroup appeared approximately 35,000-40,000 years bp in a region east of the Aral sea; subsequent branches spread to North Asia (N) and another branch (O) to South Asia via North India
Dates such as these prove that what you have just written is bull. The first amh do not enter Western eurasia until 40kya , and they do not reach eastern Europe and the levant until 34kya. Up until this time Neandethals were the dominant group in Europe. Thusly, to claim the origination of IJ and NO in eastern Europe prior to amh reaching these locations is not supported by archaeological or skeletal evidence.
As a result, if these haplogroups existed 45kya they had to have originated in africa and taken to Europe by African people.
.
Posted by the lioness, (Member # 17353) on :
quote:Originally posted by Clyde Winters:
quote:Originally posted by xyyman: Can someone explain this? I am not sure what they are trying to say here...
--- The IJ haplogroup characterizes part of the second wave of emigration from Africa that occurred via the Middle East 45,000 years bp and defines two branches I and J that emigrated northwards and eastwards into Europe. The J branch subsequently split again and contributed to the current North African population. The NO haplogroup appeared approximately 35,000-40,000 years bp in a region east of the Aral sea; subsequent branches spread to North Asia (N) and another branch (O) to South Asia via North India
Dates such as these prove that what you have just written is bull. The first amh do not enter Western eurasia until 40kya , and they do not reach eastern Europe and the levant until 34kya. Up until this time Neandethals were the dominant group in Europe. Thusly, to claim the origination of IJ and NO in eastern Europe prior to amh reaching these locations is not supported by archaeological or skeletal evidence.
As a result, if these haplogroups existed 45kya they had to have originated in africa and taken to Europe by African people.
.
no example of a Haplogroup IJ* Y-chromosome has been found among any modern human population; the existence of the Haplogroup IJ node has been inferred from the fact that certain mutations are shared in common among all Y-chromosomes belonging to the descendant haplogroups I and J. The lack of any examples of Haplogroup IJ* belonging to neither Haplogroup I nor Haplogroup J complicates any attempt to deduce the geographical location where Haplogroup IJ first appeared; however, the fact that both Haplogroup I and Haplogroup J are found among modern populations of the Caucasus, Anatolia, and Southwest Asia tends to support the hypothesis that Haplogroup IJ derived from Haplogroup F in the vicinity of West Asia or the Middle East and subsequently spread throughout Western Eurasia.
Haplogroup IJ is a descendant branch of the greater Haplogroup F (M89, P14, M213).
Posted by the lioness, (Member # 17353) on :
Stick to E though, thread topic
Posted by xyyman (Member # 13597) on :
That's my point. It does not make sense. BTW. I did not write it. This is from the ISOGG website. It seems like they are waffling.
quote:Originally posted by Clyde Winters:
quote:Originally posted by xyyman: Can someone explain this? I am not sure what they are trying to say here...
--- The IJ haplogroup characterizes part of the second wave of emigration from Africa that occurred via the Middle East 45,000 years bp and defines two branches I and J that emigrated northwards and eastwards into Europe. The J branch subsequently split again and contributed to the current North African population. The NO haplogroup appeared approximately 35,000-40,000 years bp in a region east of the Aral sea; subsequent branches spread to North Asia (N) and another branch (O) to South Asia via North India
Dates such as these prove that what you have just written is bull. The first amh do not enter Western eurasia until 40kya , and they do not reach eastern Europe and the levant until 34kya. Up until this time Neandethals were the dominant group in Europe. Thusly, to claim the origination of IJ and NO in eastern Europe prior to amh reaching these locations is not supported by archaeological or skeletal evidence.
As a result, if these haplogroups existed 45kya they had to have originated in africa and taken to Europe by African people.
.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
This is very interesting. Because, it most probably means that at one point all Niger-Congo, Cushitic and Chadic speakers were united, thus were in the same geographical location and they all spoke the same language. The whatever language spoken by the E-P2 carrier at that far away time.
So they were at the same geographical location and spoke the same language (most probably).
This by itself is interesting. It has definitive historical/archaeological ramifications.
If you want to push it a bit more. You can see how this fits with Obenga's classification of African languages exposed in the book called Origine commune de l'egyptien ancien, du copte et des langues negro-africaines modernes: Introduction a la linguistique historique africaine.
In fact, we already got the Niger-Congo, Cushitic and Chadic speakers here, all descended from the same language. Obenga's call it the Negro-Egyptian language. The only language family left is the Nilo-Saharan one. Rarely populations are formed by only one haplogroup (which can later drift). Obenga's of course already exposed the linguistic basis for including Nilo-Saharan in the Negro-egyptian language family. But other linguists, like Blench, also combines the Niger-Congo and Nilo-Saharan family into one. They call it "Niger-Saharan" "Kongo-Saharan" language family. So there we are, the Obenga's classification of African language family supported by genetics and other recent linguists (for the Niger-Saharan part).
This is another brick to add to the common origin of African people (postdating the main OOA migration). For example, something like the headrests, could be a physical representation of this common origin. On the most basic level, the fact that African populations usually looks like each other in a rough manner is another clue (in the sense that Europeans and Asians looks like each other too in a rough manner). If African populations would have been isolated from each other for a long time (let's say from before the OOA). There's no reason why they wouldn't have changed physical appearance like non-African poplations did when they left the continent during the OOA migration.
Also, while admixture and the OOA bottleneck can explain part of the situation. It doesn't seem enough to explain how come African people (as other people like Europeans, Asians, Native Americans) cluster up with each other on genetic distance calculations. We can understand why Europeans, Asians and Native Americans populations cluster up with each others respectively. For one, they have their common origin in the main OOA migration. And they also have other period of (multiple) common origin to colonize Europe, East Asia and America respectively. As any people close geographically they also admixed with each others, so we understand why they cluster up with each others in term of genetic distance. For this to work with African populations, like Europeans, Asians and Native Americans, they also must have a common origin post-dating the main OOA migration. And this is what I just noted in this thread both genetically and linguistically.
Posted by typeZeiss (Member # 18859) on :
quote:Originally posted by xyyman: Nice synopsis TP.
@ Lioness. So...We are back to White women leaving their white men behind...20ya...in search of black men. Or were they hunting in Africa for food and decided to stay and not go back?
I know some brothas like to think they are all that but...
They were enslaved and brought to North and West africa where they would then become concubines.
quote:Originally posted by Amun-Ra The Ultimate: This is very interesting. Because, it most probably means that at one point all Niger-Congo, Cushitic and Chadic speakers were united, thus were in the same geographical location and they all spoke the same language. The whatever language spoken by the E-P2 carrier at that far away time.
So they were at the same geographical location and spoke the same language (most probably).
This by itself is interesting. It has definitive historical/archaeological ramifications.
If you want to push it a bit more. You can see how this fits with Obenga's classification of African languages exposed in the book called Origine commune de l'egyptien ancien, du copte et des langues negro-africaines modernes: Introduction a la linguistique historique africaine.
In fact, we already got the Niger-Congo, Cushitic and Chadic speakers here, all descended from the same language. Obenga's call it the Negro-Egyptian language. The only language family left is the Nilo-Saharan one. Rarely populations are formed by only one haplogroup (which can later drift). Obenga's of course already exposed the linguistic basis for including Nilo-Saharan in the Negro-egyptian language family. But other linguists, like Blench, also combines the Niger-Congo and Nilo-Saharan family into one. They call it "Niger-Saharan" "Kongo-Saharan" language family. So there we are, the Obenga's classification of African language family supported by genetics and other recent linguists (for the Niger-Saharan part).
This is another brick to add to the common origin of African people (postdating the main OOA migration). For example, something like the headrests, could be a physical representation of this common origin. On the most basic level, the fact that African populations usually looks like each other in a rough manner is another clue (in the sense that Europeans and Asians looks like each other too in a rough manner). If African populations would have been isolated from each other for a long time (let's say from before the OOA). There's no reason why they wouldn't have changed physical appearance like non-African poplations did when they left the continent during the OOA migration.
Also, while admixture and the OOA bottleneck can explain part of the situation. It doesn't seem enough to explain how come African people (as other people like Europeans, Asians, Native Americans) cluster up with each other on genetic distance calculations. We can understand why Europeans, Asians and Native Americans populations cluster up with each others respectively. For one, they have their common origin in the main OOA migration. And they also have other period of (multiple) common origin to colonize Europe, East Asia and America respectively. As any people close geographically they also admixed with each others, so we understand why they cluster up with each others in term of genetic distance. For this to work with African populations, like Europeans, Asians and Native Americans, they also must have a common origin post-dating the main OOA migration. And this is what I just noted in this thread both genetically and linguistically.
The Sahara was once green, turns dry in the east around 4,000 BC a little later in the west. So probably during the green phase everyone was in one geographical location
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote:Originally posted by typeZeiss:
quote:Originally posted by Amun-Ra The Ultimate: This is very interesting. Because, it most probably means that at one point all Niger-Congo, Cushitic and Chadic speakers were united, thus were in the same geographical location and they all spoke the same language. The whatever language spoken by the E-P2 carrier at that far away time.
So they were at the same geographical location and spoke the same language (most probably).
This by itself is interesting. It has definitive historical/archaeological ramifications.
If you want to push it a bit more. You can see how this fits with Obenga's classification of African languages exposed in the book called Origine commune de l'egyptien ancien, du copte et des langues negro-africaines modernes: Introduction a la linguistique historique africaine.
In fact, we already got the Niger-Congo, Cushitic and Chadic speakers here, all descended from the same language. Obenga's call it the Negro-Egyptian language. The only language family left is the Nilo-Saharan one. Rarely populations are formed by only one haplogroup (which can later drift). Obenga's of course already exposed the linguistic basis for including Nilo-Saharan in the Negro-egyptian language family. But other linguists, like Blench, also combines the Niger-Congo and Nilo-Saharan family into one. They call it "Niger-Saharan" "Kongo-Saharan" language family. So there we are, the Obenga's classification of African language family supported by genetics and other recent linguists (for the Niger-Saharan part).
This is another brick to add to the common origin of African people (postdating the main OOA migration). For example, something like the headrests, could be a physical representation of this common origin. On the most basic level, the fact that African populations usually looks like each other in a rough manner is another clue (in the sense that Europeans and Asians looks like each other too in a rough manner). If African populations would have been isolated from each other for a long time (let's say from before the OOA). There's no reason why they wouldn't have changed physical appearance like non-African poplations did when they left the continent during the OOA migration.
Also, while admixture and the OOA bottleneck can explain part of the situation. It doesn't seem enough to explain how come African people (as other people like Europeans, Asians, Native Americans) cluster up with each other on genetic distance calculations. We can understand why Europeans, Asians and Native Americans populations cluster up with each others respectively. For one, they have their common origin in the main OOA migration. And they also have other period of (multiple) common origin to colonize Europe, East Asia and America respectively. As any people close geographically they also admixed with each others, so we understand why they cluster up with each others in term of genetic distance. For this to work with African populations, like Europeans, Asians and Native Americans, they also must have a common origin post-dating the main OOA migration. And this is what I just noted in this thread both genetically and linguistically.
The Sahara was once green, turns dry in the east around 4,000 BC a little later in the west. So probably during the green phase everyone was in one geographical location
Personally, I view the Green Sahara phase as an admixture phase. The Bantu migration is another admixture phase for example. Let's recall that we're only in the years 2000s and the Green Sahara lasted more than 3000 years. A lot of time to move around and admix with each others.
If you take into account that most African language families and the E-P2 haplogroup originate in East Africa (post dating the main OOA migration) then I would say that Eastern Africa, around Sudan maybe, is the geographical location of this common origin of African people.
Posted by Trollkillah # Ish Gebor (Member # 18264) on :
quote:Originally posted by Amun-Ra The Ultimate: E-P2 (PN2) is one of the main haplogroup in Africa along with other E, A and B haplogroups.
It has its origin after the main OOA migration of non-Africans. E-P2 carriers weren't part of the main OOA migration.
So E-P2 unites African people and is probably one of the main haplogroup among Ancient Egyptians along with other A, B and E haplogroups.
Most Niger-Congo(Kordofanian), Cushitic and Chadic speakers are carriers of E-P2.
quote: Tanzania is the only region of Africa where populations speak languages classified as belonging to the 4 major language families present in Africa: Afro-Asiatic, Nilo-Saharan, Niger-Kordofanian, and Khoisan (Greenberg 1963). The Hadza and Sandawe, who speak a click language classified as Khoisan, are thought to be indigenous to Tanzania. However, populations speaking languages belonging to the other 3 language families are thought to have migrated into Tanzania from the Sudan (Nilotic Nilo-Saharan speakers), Ethiopia (Cushitic Afro-Asiatic speakers), and West Africa (Bantu Niger-Kordofanian speakers) within the past 5,000 years (Ambrose 1982; Newman 1995).
--Sarah A. Tishkoff
Evolutionary history of mtDNA haplogroup structure in African populations inferred from mtDNA d-loop and RFLP analysis.
quote:Originally posted by xyyman: Nice synopsis TP.
@ Lioness. So...We are back to White women leaving their white men behind...20ya...in search of black men. Or were they hunting in Africa for food and decided to stay and not go back?
I know some brothas like to think they are all that but...
They were enslaved and brought to North and West africa where they would then become concubines.
People can be related genectically and not have the same origins, as a result of admixture. Below are 18 points that support a Saharan--not East African origin of the Niger-Congo speakers.
1. The Saharan population hunted animals with the bow-and arrow; they are associated with the Ounanian culture. The Ounanian culture existed 12kya [2].
2. 10,000 BC there was continuity between the populations in the Maghreb and southern Sahara referred to as Capsians, Iberomaurusians, and Mechtoids [3].
3. The Ounanian culture is associated with sites in central Egypt, Algeria, Mali, Mauretania and Niger. There are no East African sites. . 4. The original homeland of the Niger-Congo speakers was probably situated in the Saharan Highlands.
5. Proto-Niger- Congo people developed an agro-pastoral economy which included the cultivation of millet, and domestication of cattle (and sheep).
6. The Niger-Congo speakers probably began to exit the Saharan Highlands during the Ounanian period. By the 8th millennium BC Saharan-Sudanese pottery was used in the Air [22]. Ceramics of this style have also been found at sites in the Hoggar [22-23]. Dotted wavy-line pottery has also been discovered in the Libyan Sahara [22]. Again no sites are found in East Africa.
7. They migrated from the Highlands into Nubia.
7a. Genetic evidence supports the upper Nile settlement for the Niger-Congo speakers. Rosa et al, in a paper discussing the y-Chromosomal diversity in the population of Guinea-Bissau, noted that while most Mande & Balanta carry the E3a-M2 gene, there are a number of Felupe-Djola, Papel, Fulbe and Mande carry the M3b*-M35 gene the same as many non-Niger-Congo speaking people in the Sudan.
8. They were the C-Group people.
9. Researchers have conclusively proven that the Dravidians are related to the Niger-Congo speaking group and they originally lived in Nubia [7]. The Dravidians and C-Group people of Nubia used 1) a common BRW [7]; 2) a common burial complex incorporating megaliths and circular rock enclosures [7] and 3) a common type of rock cut sepulcher [7] and writing system [50-51].
10. The BRW industry diffused from Nubia, across West Asia into Rajastan, and thence to East Central and South India [30]. Singh [30] made it clear that he believes that the BRW radiated from Nubia through Mesopotamia and Iran southward into India
11. The mtDNA haplogroups L1, L2, L3 and U5 are associated with Niger-Congo speakers. Phylogenetically all the Eurasian mtDNA branches descend from L3. The Pan-African haplotypes are 16189,16192,16223, 16278,16294, 16309, qnd 16390. This sequence is found in the L2a1 haplotype which is highly frequent among the Mande speaking group and the Wolof.
12. The phylogeography of y-Chromosome haplotypes shared among the Niger-Congo speakers include A,B, Elb1a, E1b1b, E2, E3a and R1 [57] (See: Figures 1-2). The predominate y-Chromosome among the Niger-Congo is M2, M35, and M33.
Haplogroup E has three branches carried by Niger-Congo populations E1, E2 and E3. The E1 and E2 clines are found exclusively in Africa. Haplogroup E3 is also found in Eurasia. Haplogroup E3 subclades are E3b, E-M78, E-M81 and E-M34. The E clades probably originated in Saharan Africa. This is based on the fact that the Niger-Congo people carry this haplogroup at high frequencies.
The majority of Niger-Congo speakers belong to E1b1a, Elb1b, E2 and R1. Around 90% belong to y-Chromosome group E (215,M35*).
Y-Chromosome haplogroup A is represented among Niger-Congo speakers. In West Africa, under 5% of the NC speakers belong to group A. Most Niger-Congo speakers who belong to group A are found in East Africa and belong to A3b2-M13: Kenya (13.8) and Tanzanian (7.0%).
13. The Bantu originated in Saharan Africa not East Africa. The Bantu expansion is usually associated with the spread of y-Chromosome E3a-M2. The most common branch of the V-38 haplogroup is E-M2. E-M2 dates to around 25ky old. It probably originated in the Highland area during the Ounanian period.
14. Some researchers claim that: The downstreams SNP E-M180 possibly originated on the moist south-central Saharan savannah/grassland of northern West Africa during the early Holocene period. Much of the population that carried E-M2 retreated to southern West Africa with the drying of the Sahara. These later people migrated from Southeastern Nigeria and Cameroon ~8.0 kya to Central Africa, East Africa, and Southern Africa causing or following the Bantu expansion.[4][5][6] According to Wood et al. (2005) and Rosa et al. (2007), such population movements from West Africa changed the pre-existing population Y chromosomal diversity in Western, Central, Southern and southern East Africa, replacing the previous haplogroups frequencies in these areas with the now dominant E1b1a1 lineages. See: http://en.wikipedia.org/wiki/Haplogroup_E-V38 In Kenya the frequentcy for E3a-M2 is 52%; and 42% in Tanzania. In Burkina Faso high frequentcies of E-M2* and E-M191* are also represented. It is interesting to note that among the Mande speaking Bisa and Mandekan there are high frequentcies of E-M2*. This is in sharp contrast to the Marka and South Samo who have high frequencies of E-M33.
15. The pristine form of R1-M173 is found in Africa. Y-Chromosome R is characterized by M207/ V45. The V45 mutation is found among NC speakers. The R1b mutations include V7, V8, V45, V69 and V88. The frequentcy of R1-M173 varies among Niger-Congo speakers. The frequentcy of R-M173 range between 3-54%. The most frequent subtype in Africa is V88 (R1b1a). Haplogroup R1b1a ranges between 2-20% among the Bantu speakers.The highest frequentcy of R1 is found among Fulbe or Fulani speakers
16. First use Saharo-Sudanese Pottery in Niger 10,000BC
17. First use of cereals in the Sahara 9400 BC
18.7500 BC domestication of sheep and goats in Sahara.
.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote:Originally posted by Clyde Winters: [QB] People can be related genectically and not have the same origins, as a result of admixture.
Every human populations (as well as individuals for that matter) are the results of admixtures. When I'm talking about the common origin of people (like Niger-Kordofanian, Cushitic and Chadic speakers), I'm talking about the common origin of the greater part of their ancestry. In this case, the greater part of their genetic ancestry.
For example, Yoruba are over 90% from the P2 haplogroups (P2/e1b1a). For Somali it's over 80% (P2/e1b1b) of their population.
90% and 80% are really large numbers. So it constitute the greater part of those populations (NK, Cushitic, Chadic) ancestry on the Y-DNA side.
The MtDNA counterparts of E-P2 are L2a, L3bf, L3cd, L3eikx, L0a and possibly a few other haplogroups. Those are the MtDNA haplogroups spread across all E-P2 populations in Africa, not just limited to one region or a few E-P2 populations.
For example, Yoruba got 75.75% and Somali 66.93% of L2a, L3bf, L3cd, L3eikx, L0a1 (using numbers from the Hirbo study linked below).
Some haplogroups are not shared between Yoruba, Somali and various E-P2 populations like L4 or M and N haplogroups. Those are the products of more recent admixtures between E-P2 migrants and other populations. Some MtDNA haplogroups are shared across many E-P2 populations but not all of them (or not almost all of them), or not across all regions where the E-P2 lineage is present in large proportion, so I didn't include those haplogroups as part of the original E-P2 population (even if it could be possible due to genetic drift in extent population).
The idea is to determine the mtDNA counterparts of the E-P2 lineages as it spread all across Africa. All regions, almost all populations. So those mtdNA counterparts must be present in almost ALL populations with a high level of E-P2. Only the migrations of E-P2 lineage carriers across all Africa (which did happen for sure) can explain the pan-African widespread nature of those MtDNA haplogroups.
As a side note, the origin of an haplogroup doesn't matter. What is important here is that it eventually found its way into the E-P2 populations before the E-P2 population split and migrated toward different directions in Africa (eventually turning into E1b1a for West Africa and E1b1b for East Africa). Also it doesn't really matter if E-P2 originated in the Northeastern Africa or anywhere between Chad and the Nile, or anywhere else, although basic logic would indicate that it has a greater chance of having originated where there's a greater diversity and frequency of E-P2. That's why biologist locate the origin of E-P2 in Eastern Africa.
quote:Originally posted by Clyde Winters: [QB] The E clades probably originated in Saharan Africa.This is based on the fact that the Niger-Congo people carry this haplogroup at high frequencies.[qb]
It's true that Niger-Congo(Niger-Kordofanian) speakers carry this haplogroup in great frequencies but so does Cushitic and Chadic speakers. Northeastern Africa also has a greater diversity of the E haplogroup.
The Y-DNA and mtDNA haplogroup frequencies for Somali (Cushitic speakers) and Yoruba (Niger-Kordofanian speakers) used in this post, as well as other African populations, were taken from the Hirbo study (starting at Appendix 6a ii, p195): http://drum.lib.umd.edu/handle/1903/11443 Posted by Clyde Winters (Member # 10129) on :
quote:Originally posted by Amun-Ra The Ultimate:
quote:Originally posted by Clyde Winters: [QB] People can be related genectically and not have the same origins, as a result of admixture.
Every human populations (as well as individuals for that matter) are the results of admixtures. When I'm talking about the common origin of people (like Niger-Kordofanian, Cushitic and Chadic speakers), I'm talking about the common origin of the greater part of their ancestry. In this case, the greater part of their genetic ancestry.
For example, Yoruba are over 90% from the P2 haplogroups (P2/e1b1a). For Somali it's over 80% (P2/e1b1b) of their population.
90% and 80% are really large numbers. So it constitute the greater part of those populations (NK, Cushitic, Chadic) ancestry on the Y-DNA side.
The MtDNA counterparts of E-P2 are L2a, L3bf, L3cd, L3eikx, L0a and possibly a few other haplogroups. Those are the MtDNA haplogroups spread across all E-P2 populations in Africa, not just limited to one region or a few E-P2 populations.
For example, Yoruba got 75.75% and Somali 66.93% of L2a, L3bf, L3cd, L3eikx, L0a1 (using numbers from the Hirbo study linked below).
Some haplogroups are not shared between Yoruba, Somali and various E-P2 populations like L4 or M and N haplogroups. Those are the products of more recent admixtures between E-P2 migrants and other populations. Some MtDNA haplogroups are shared across many E-P2 populations but not all of them (or not almost all of them), or not across all regions where the E-P2 lineage is present in large proportion, so I didn't include those haplogroups as part of the original E-P2 population (even if it could be possible due to genetic drift in extent population).
The idea is to determine the mtDNA counterparts of the E-P2 lineages as it spread all across Africa. All regions, almost all populations. So those mtdNA counterparts must be present in almost ALL populations with a high level of E-P2. Only the migrations of E-P2 lineage carriers across all Africa (which did happen for sure) can explain the pan-African widespread nature of those MtDNA haplogroups.
As a side note, the origin of an haplogroup doesn't matter. What is important here is that it eventually found its way into the E-P2 populations before the E-P2 population split and migrated toward different directions in Africa (eventually turning into E1b1a for West Africa and E1b1b for East Africa). Also it doesn't really matter if E-P2 originated in the Northeastern Africa or anywhere between Chad and the Nile, or anywhere else, although basic logic would indicate that it has a greater chance of having originated where there's a greater diversity and frequency of E-P2. That's why biologist locate the origin of E-P2 in Eastern Africa.
quote:Originally posted by Clyde Winters: [QB] The E clades probably originated in Saharan Africa.This is based on the fact that the Niger-Congo people carry this haplogroup at high frequencies.[qb]
It's true that Niger-Congo(Niger-Kordofanian) speakers carry this haplogroup in great frequencies but so does Cushitic and Chadic speakers. Northeastern Africa also has a greater diversity of the E haplogroup.
The Y-DNA and mtDNA haplogroup frequencies for Somali (Cushitic speakers) and Yoruba (Niger-Kordofanian speakers) used in this post, as well as other African populations, were taken from the Hirbo study (starting at Appendix 6a ii, p195): http://drum.lib.umd.edu/handle/1903/11443
None of this proves that modern African groups originated Northeast Africa.
.
Posted by beyoku (Member # 14524) on :
quote:Originally posted by Amun-Ra The Ultimate:
quote:Originally posted by Clyde Winters: [QB] People can be related genectically and not have the same origins, as a result of admixture.
Every human populations (as well as individuals for that matter) are the results of admixtures. When I'm talking about the common origin of people (like Niger-Kordofanian, Cushitic and Chadic speakers), I'm talking about the common origin of the greater part of their ancestry. In this case, the greater part of their genetic ancestry.
For example, Yoruba are over 90% from the P2 haplogroups (P2/e1b1a). For Somali it's over 80% (P2/e1b1b) of their population.
90% and 80% are really large numbers. So it constitute the greater part of those populations (NK, Cushitic, Chadic) ancestry on the Y-DNA side.
This is patently False. What does "ancestry on the Y-DNA side" even mean? They Y-dna does not code for autosomal dna regardless of you thinking it does. You are only looking at 30 generations. You are speaking on the ancestry of 30 MEN in the image below but you ignore the other 1,073,741,794 !
What number is larger you ding bat 30 or 1,073,741,794? Man GTFOOHWTBS ! Posted by Amun-Ra The Ultimate (Member # 20039) on :
@ Clyde Winters
You can disagree with it or not but most biologists locate the origin of the E (M96) haplogroup in Northeastern Africa. Same for the related downstream E-P2 haplogroup (see above).
Posted by beyoku (Member # 14524) on :
quote:Originally posted by Amun-Ra The Ultimate: @ Clyde Winters
You can disagree with it or not but most biologists locate the origin of the E (M96) haplogroup in Northeastern Africa. Same for the related downstream E-P2 haplogroup (see above).
Why Would it be East African when its Ancestor DE* is firmly found in West Africa? Also most....not all but MOST of the E* lineages are western African? The oldest lineage of E........E1a = West African...what about E2...That looks to be West African as well.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
@beyoku Stop being a stupid racist as well as exhibiting ridiculous knee jerk reactions everybody can see. Your post comes down to not being able to use uniparental haplogroups at all to analyze population movements and migrations, which is part of their very purpose and are used in many scientific study for such purpose. There's also other purpose like medical, etc.
To analyse population movements and migrations you must use uniparental as well as autosomal DNA (as well as other linguistic and archaeological data). This is what I do in this linked post (as well as various posts on this site):
You may have 1,073,741,794 ancestors but if almost over 90% of them derive from the same Y-DNA haplogroup (and the same MtDNA haplogroups) it means those ancestors share a lot of autosomal DNA between each others. Those 1,073,741,794 ancestors are related to each others through the E-P2 and various MtDNA haplogroups in great proportion.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote:Originally posted by beyoku:
quote:Originally posted by Amun-Ra The Ultimate: @ Clyde Winters
You can disagree with it or not but most biologists locate the origin of the E (M96) haplogroup in Northeastern Africa. Same for the related downstream E-P2 haplogroup (see above).
Why Would it be East African when its Ancestor DE* is firmly found in West Africa? Also most....not all but MOST of the E* lineages are western African? The oldest lineage of E........E1a = West African...what about E2...That looks to be West African as well.
You can disagree with it or not but most biologists locate the origin of the E (M96) haplogroup in Northeastern Africa. Same thing for E-P2:
quote: Using the principle of the phylogeographic parsimony, the resolution of the E1b1b trifurcation in favor of a common ancestor of E-M2 and E-M329 strongly supports the hypothesis that haplogroup E1b1 originated in eastern Africa , as previously suggested [10], and that chromosomes E-M2 , so frequently observed in sub-Saharan Africa, trace their descent to a common ancestor present in eastern Africa .
-- from A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms (Trombetta 2011)
Posted by beyoku (Member # 14524) on :
quote:Originally posted by Amun-Ra The Ultimate: @beyoku Stop being a stupid racist as well as exhibiting ridiculous knee jerk reactions everybody can see. Your post comes down to not being able to use uniparental haplogroups at all to analyze population movements and migrations, which is part of their very purpose and are used in many scientific study for such purpose. There's also other purpose like medical, etc.
To analyse population movements and migrations you must use uniparental as well as autosomal DNA (as well as other linguistic and archaeological data). This is what I do in this linked post (as well as various posts on this site):
You may have 1,073,741,794 ancestors but if almost over 90% of them derive from the same Y-DNA haplogroup (and the same MtDNA haplogroups) it means those ancestors share a lot of autosomal DNA between each others. Those 1,073,741,794 ancestors are related to each others through the E-P2 and various MtDNA haplogroups in great proportion.
Wrong again dumbo. 1 - Please detail the data they can pull from Y-dna studies? Is this something you can do without googling? 2 - Please explain the very different Autosomal makeup of Mandinka and Tutsi..........both of which are about 80% E1b1a. Please explain the very different autosomal makeup of Tunisian Berbers and Ethiopians groups again who have E1b1b lineages that reach fixation.
Here is a bonus question. While your are at it please explain the differences between Greeks/Albanians and Chad Basin populations? Interesting BOTH population have similiar frequencies of E1b1 AND R1b1. What about African Oludeme and Irish/Portugese....all are both Very high in R1b1 80-90%.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
Stop acting like a dumbass and answer some questions yourself: - Please explain why are you so butthurt that Niger-Congo, Cushitic and Chadic speakers are united by E-P2 and various MtDNA lineages? - Please explain why it hurts you so bad that East and West Africans share common lineages and were one people after the OOA migrations of non-Africans? - Please explain why you feel the need to come here on this forum out of your way to try to deny this clear past unity and shared history between Niger-Congo, Cushitic and Chadic speakers?
Posted by Swenet (Member # 17303) on :
Please explain why not just any person, but the empirical evidence is staring you in the face and telling you that you're full of bs when you say that the West Africans you refer to split only recently, during the Green Sahara (using not any scientific reference, but your own non-existent grasp of population genetics):
quote:We date the initial split between Yoruba and out of African populations to 90,000-100,000 years ago with substantial gene flow occurring until nearly 50,000 years ago, and obtain congruent results with the autosomes and the X chromosome.
Damn! You just took a haymaker to the face from Song et al! All that "Africans were one people until 3000 years ago" crap... just give it up, cesspool mouth. The stench of bs emanating from your propagandistic posts is unbearable at this point.
Not unimportant in light of the propaganda you're evangelizing in your fraudulent threads; apparently the software of Song et al has no problem detecting the migration of L3 lineages ~50kya towards West/ Central African aborigines, using ancestral Europeans and Indians as models! Guess where that leaves your claim that a high diversity and frequency of L3 in certain East Africans doesn't imply a closer genetic affinity between them and OOA populations.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote:Originally posted by Swenet: Please explain why not just any person, but the empirical evidence is staring you in the face and telling you that you're full of bs when you say that the West Africans you refer to split only recently, during the Green Sahara (using not any scientific reference, but your own non-existent grasp of population genetics):
You're lying to us again. If you need to lie about my position, that you know otherwise very well, to try to get the upper hand, it means your position is very weak.
Dates are never precise in genetic studies because they are always estimation based on a fixed mutation rates of change like glottochronology.
I locate the common origin of East and West Africans between the OOA migrations and the foundation of Ancient Egypt. The exact time would be the time the E-P2 population split (first within the Northeastern African regions) and migrated toward their respective contemporary regions (first toward the Sahara, then West Africa and eventually toward southern Africa with the Bantu migrations) along with their MtDNA female counterparts. Nobody knows the exact time since it would always be an estimation unless there's ancient DNA (carbon dated). But suffice to say it's AFTER the OOA migrants left and separated from their African brethren since E-P2 and the various African L3 MtDNA haplogroups (L3eikx, L3bf, L3cd, etc) shared between African people (East, West, South) are downstream(after) the OOA L3 and CT split.
quote:
quote:We date the initial split between Yoruba and out of African populations to 90,000-100,000 years ago with substantial gene flow occurring until nearly 50,000 years ago, and obtain congruent results with the autosomes and the X chromosome.
Damn! You just took a haymaker to the face from Song et al! All that "Africans were one people until 3000 years ago" crap... just give it up, cesspool mouth. The stench of bs emanating from your propagandistic posts is unbearable at this point.
Not unimportant in light of the propaganda you're evangelizing in your fraudulent threads; apparently the software of Song et al has no problem detecting the migration of L3 lineages ~50kya towards West/ Central African aborigines, using ancestral Europeans and Indians as models! Guess where that leaves your claim that a high diversity and frequency of L3 in certain East Africans doesn't imply a closer genetic affinity between them and OOA populations.
You're so desperate to try to deny the obvious common origin of East and West Africans that you use a study that makes you shoot yourself in the foot.
The study you try to use also mentions and I quote: " substantial gene flow occurring until nearly 50,000 years ago". Of course the date as all the analysis are an estimation (this study is model based) but this substantial gene flow is what represent the greater part of their ancestry. Of course when modern African migrated to their respective regions they met small groups of hunter gatherers but due to demographic expansion/technological advantage those hunter gatherers are not there anymore in West Africa as such and are only present in small proportion as Khoisan and Aka-Mbuti (A, B haplogroups) people in the rest of Africa.
For example, Yoruba got 92% of haplogroup P2/E1b1a, they share with East Africans like Somali, and only 8% of B-M60. This 8% represent what you call the aboriginal component to West Africans. Clearly it doesn't form the greater part of their ancestry. This post-OOA migrations is what has the larger historical and cultural ramifications.
I mentions the Green Saharan connection to modern West Africans (which are post OOA migrants for the majority of their population/genome) here:
When modern West Africans arrived at their current West African regions after their migrations from the desiccating Green Sahara. They met there small groups of hunter-gatherers from the B haplogroup (probably A hg too). Which were then probably completely assimilated into the Green Saharan West African migrants populations since no cultural traces (like languages or lifestyles) of those people are left in that region. The greater part of West African ancestry are shared with the various E-P2 populations (NK, Cushitic, Chadic speakers) in Africa. For Yoruba it's 92% of P2/e1b1a and 75.75% of various MtDNA haplogroups shared with other E-P2 populations like Somali.
Swenet I got some questions for you: - Why are you so desperate to deny the obvious common origin of modern African people (East, West, South) after the OOA migrations? - Why are you so butthurt and dogmatic about it? Does it changes something in your life? Why do you feel you need to deny the obvious then?
Posted by Tukuler (Member # 19944) on :
There weren't a billion people on earth in the 1300's and needful to say all of them can't be ancestral to every single individual. It's not a simple mathematics projection. What's left unconsidered is the dampering out effect of overlap in "parentage" (cousin marriage, marriage to distant relatives who didn't know they were related, etc.).
Nonetheless straight line maternal mtDNA and paternal nrY chromosome lineages don't tell nearly as much about an individual's or even whole populations' geographic geneaology as do the autosomes because the sex linked genetic indicators, outside of the direct maternal and paternal ones get lost each generation.
Posted by beyoku (Member # 14524) on :
quote:Originally posted by Tukuler:
Nonetheless straight line maternal mtDNA and paternal nrY chromosome lineages don't tell nearly as much about an individual's geographic geneaology as the autosomes because the sex linked genetic indicators outside of the straight maternal and paternal ones are lost each generation compone
This. And he has a hard time understanding this. that said in reference to the math you have to factor in death. The numbers are correct.
Posted by Swenet (Member # 17303) on :
quote:Originally posted by Amun-Ra The Ultimate: You're lying to us again. If you need to lie about my position
I'm lying? Don't make me pull out your weirdo posts where you claim that all speakers of Negro-Egyptian genetically coalesce to a MRCA with the proto speakers of this language family, somewhere in the terminal pleistocene. You also cited Ehret to this effect where he says that most modern African language phyla were relatively recently in East Africa. Your posts are on record. No use in flip flopping now, liar.
quote: Originally posted by Amun-Ra The Ultimate: I locate the common origin of East and West Africans between the OOA migrations and the foundation of Ancient Egypt.
I know you do. That's what why Song et al 2014 just buried the sloppy propagandistic scheme you try to evangelize here, but, to no avail.
quote: Originally posted by Amun-Ra The Ultimate: For example, Yoruba got 92% of haplogroup P2/E1b1a, they share with East Africans like Somali, and only 8% of B-M60. This 8% represent what you call the aboriginal component to West Africans. Clearly it doesn't form the greater part of their ancestry.
This crappy statement presumes that haplogroup profiles are stable throughout time and accurate portrayals of all the genetic history of a population. Are they? Is E1b1a the only major Y-DNA expansion that impacted the ancestral Yoruba since West Africa was settled by AMHs? Post evidence.
^Watch the clown cop-out again
Posted by Tukuler (Member # 19944) on :
The numbers are mathamatically correct as far 2 to the x power.
The fact remains that 30 generations ago there were not a billion people on earth hence no one had a billion ancestors in the 13th century, just research projected global population figures or write and ask your favorite geneticist of choice to explain how to sync number of generations to actual population sizes or just note how ridiculously astronomical the figure is at 100 generations ago and what that would imply.
Obviously at various points in time the generation pyramid is more like a diamond i.e. shrinking not growing.
Posted by Swenet (Member # 17303) on :
quote:Originally posted by beyoku:
quote:Originally posted by Tukuler:
Nonetheless straight line maternal mtDNA and paternal nrY chromosome lineages don't tell nearly as much about an individual's geographic geneaology as the autosomes because the sex linked genetic indicators outside of the straight maternal and paternal ones are lost each generation compone
This. And he has a hard time understanding this. that said in reference to the math you have to factor in death. The numbers are correct.
You'd have to factor in that autosomally there are no linear "lineages" and you and I, for instance, may share 10000s ancestors since slavery, just because the way human autosomal DNA is intertwined due to recombination. The ancestors in your image don't have to add up to billions of people at once.
Posted by Tukuler (Member # 19944) on :
They simply cannot add up to billions. Human population has increased over the years. The exponential number of ancestors is patently false because using it to go back to our pleistocene beginnings would require more people than has ever lived altogether through time to be alive all in one generation.
Posted by Swenet (Member # 17303) on :
Tracing one person's lineage back in time for a few generations in principle forms a binary tree of parents, grandparents, great-grandparents and so on. However, the number of individuals in such an ancestor tree grows exponentially and very soon exceeds the population from which the ancestors were drawn. A human alive today would, over 30 generations (going back to about the High Middle Ages), have 2^30 or about 1.07 billion ancestors, more than the world population at the time. Posted by Tukuler (Member # 19944) on :
Obviously if only N people were alive then N+ ancestors are impossible for the time period.
The point remains uniparentals show deep lineage for fathers fathers father way back into time and are very narrow slits compared to the wide berth of information obtainable from autosomes which have bits from all ancestors not just the 2 uniparentals indicate no matter which generation is chosen.
I can't find it but I prepped an expo tree for 3 or 4 generations of a hypothetical AfrAm male illustrating the massive loss of African genetics if only his Euro male nrY and NativeAmer mtDNA were considered. His uniparentals would make him a mestizo whereas his autosomes clearly demostrated his "Africanity."
Here is something similar, and more eye pleasing. In my chart BLUE gggrandaddy was Euro and RED ggnonna was NativeAmer and WHITE was the Afr ancestry hidden if relying solely on uniparentals.
Posted by Swenet (Member # 17303) on :
Even if one takes multiple counting into account, it's a non-sequitur that the amount of ancestors one has since the Middle Ages can't exceed the amount of people who were alive in the Middle Ages. Unless I'm missing something not yet brought to my attention in this thread, counting the amount of ancestors back in time who donated genetic material to the genepool of someone who is alive today, at some point, you're going to reach a billion, whether you count multiple times or not. That's the last thing I have to say about this axiomatic idea.
Posted by Tukuler (Member # 19944) on :
quote:Originally posted by Swenet:
Even if one takes multiple counting into account, it's a non-sequitur that the amount of ancestors since the Middle Ages can't exceed the amount of people who were alive in the Middle Ages. Unless I'm missing something not yet brought to my attention in this thread, counting the amount of ancestors back in time who donated genetic material to the genepool of someone who is alive today, at some point, you're going to reach a billion, whether you count multiple times or not. That's the last thing I have to say about this axiomatic idea.
.
No geneticists supports such an idea. At least I have not seen any genetic report even implying its correctness that the number of ones ancestors can exceed the number of people alive at any given generation. Quite to the contrary see the above posted snippets from
Ohno (1996) What prevents the exponential increase of one's ancestors?. Proc. Natl. Acad. Sci. USA Vol. 93, pp. 1527615278, December 1996
and this from the abstract
The reason that the indefinite exponential increase in the number of ones ancestors does not take place is found in the law of sibling interference, ...
Posted by Amun-Ra The Ultimate (Member # 20039) on :
To analyse population history, migrations and the common origin of Niger-Kordofanian, Cushitic and Chadic language speakers, we must use a multidisciplinary approach. On the genetic front we must use uniparental as well as autosomal DNA. In this thread, I used mainly the uniparental haplogroups. In the linked thread below, I also used autosomal, uniparental as well as archaeological and linguistic data:
Autosomally, you also can see the evidence of the common origin of NK, Cushitic and Chadic African people.
The genetic distance between various African and world populations shouldn't be a guess, or something related to our own personal prejudice and bias. The genetic distances between populations is measured in a scientific way by geneticists using genetic distance mathematical formula.
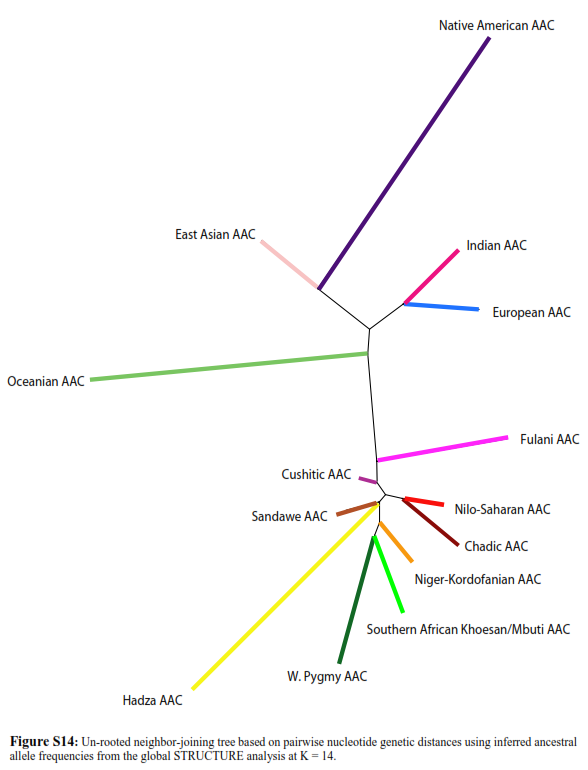
In the following genetic distance tree, using autosomal STR, we can see Cushitic, Chadic and Niger-Kordofanian speakers as well as various African populations are genetically closer to each others than they are to any non-African populations:
This graph above completely obliterate the argument of some people in this thread who try to say it was not true (autosomally) that Cushitic, Chadic and Niger-Kordofanian speakers were genetically closer to each others than toward any non-African populations. We can see that it is both true autosomally as it was with uniparental (shared E-P2 and various MtDNA haplogroups).
I could end my argumentation right here. My point is already won. That is we can clearly see that Cushitic, Chadic and Niger-Kordofanian are genetically closer to each others than they are to any Europeans or West Asian populations using autosomal DNA this time. The African AACs are at the bottom and the non-African AACs at the top of the graph.
As a side note. We know from uniparental Y-DNA and MtDNA that Fulani, for example, have some recent post-OOA Eurasian admixtures. This would explain why the Fulani AAC cluster is closer to non-Africans than for example Niger-Kordofanian and Nilo-Saharan speakers are to non-Africans. The Fulani AAC is still much closer to the Niger-Kordofanian and Nilo-Saharan AAC than to any non-African AACs.
There's many other graph showing the relatively close genetic distances between various African populations like HERE, HERE, HERE, and HERE. This is also true for Eurasians, East Asians and Native Americans populations toward each other respectively. In all those graphs, we can see African populations clustering close to each others because of 2 main aspects 1)Relatively recent common origin 2)Relatively recent admixtures. By relatively recent we mean post-OOA migrations.
But to be completely sure, we can go further than this. We must make sure Cushitic and Chadic populations are not genetically close to each others and to Niger-Kordofanian populations because of recent admixtures.
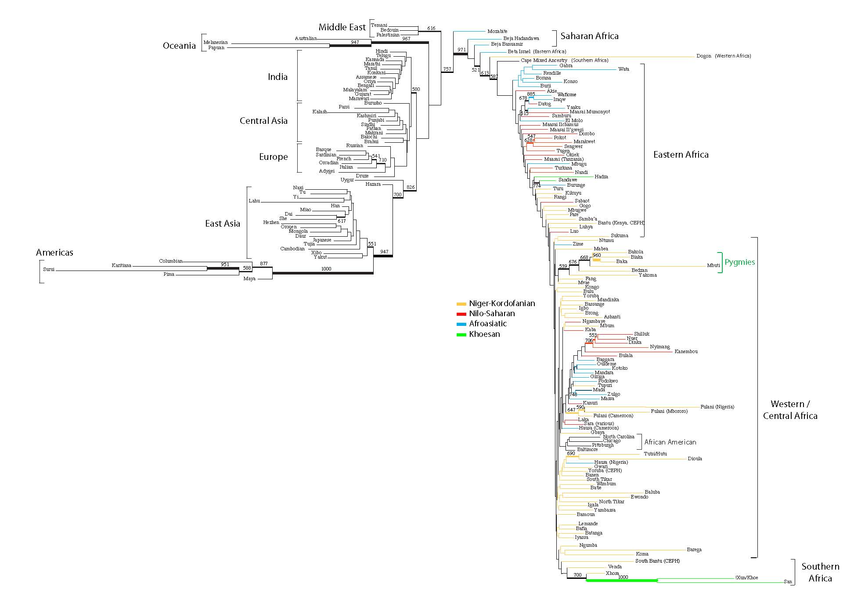
For this, lets use another genetic distance tree from the same study (Figure S7, p33). Links to bigger image below or in the study:
This above genetic distance tree using autosomal STR is also enough by itself to show African populations are genetically closer to each others than they are toward any non-African populations. This genetic distance tree is on scale, so we can measure the genetic distances with a ruler or by eye. We measure the distances by calculating the horizontal distance undertaken by traveling from one population node to another.
For example, we can see Yoruba and Mandinka are very close to each others in term of genetic distance. While Igbo are a bit further away from Yoruba but not that much. All those populations (Yoruba, Mandinka and Igbo) are clearly much closer to each others than to non-African populations like Bedouin or French which are completely on the other side.
Generally, African populations cluster in term of autosomal genetic distance on the right side of the tree, and non-African on the left side. But more precisely, it seems to be mostly the populations between the Gabra (top right) and the Venda (bottom right) which are genetically closer to each others than to Eurasian populations. African populations between Mozabite and Cape Mixed Ancestry are kind of in between African and non-African populations. Mozabite are closer to non-African populations (Bedouin), while Beja Hadandawa are just a tad closer to African populations like Yoruba than they are to the Bedouin and other non-African populations.
What is important for us is that between Gabra and Venda, all those populations are genetically closer to each others than they are to any Eurasian populations like in the Middle East, Oceania or Europe.
The next step into our investigation is to verify (for example) that Cushitic speakers like the Gabra are not recently admixed with Niger-Kordofanian speakers like Yoruba and Bantu. A bit like we did with Somali populations above.
For this, we will first use again the table of haplotype frequencies from the Hirbo study (starting at Appendix 6a ii, p195): DOWNLOAD HERE.
If you take the time to look at it, this confirms Gabra are not recently admixed with Niger-Kordofanian speakers (like Yoruba) or Bantu populations.
Gabra: - Percentage of E1b1b: 82.6%(6.9+58.6+3.4+10.3+3.4) - No E1b1a and no L3e, so no evidence of recent admixtures between the Gabra and Niger-Kordofanian speakers characterized by E1b1a and L3e. They only share the related upstream(older) basal P2 and African L3(L3eikx, etc) lineages with West Africans/Yoruba and Bantu populations as does all African populations cited in this thread (NK, Cushitic, Chadic). - It's also interesting to note that Gabra have a low level of Eurasian Y-DNA haplogroups like F descendants (6.8%) and about 54.83% (100-45.17=54.83) of Eurasian M, N descendants haplogroups. Which is typical of female mediated recent admixture (post OOA at the very least) with Semitic (ethiosemitic) speakers and Muslim Arabs speakers in Eastern Africa. The same can be seen in Somali, Afar and various Northeastern African populations. We can also see the frequencies of Eurasian M and N haplogroups in Gabra people here too.
With this I have proved my point again. Gabra, which are Cushitic speakers, NOT recently admixed with Niger-Kordofanian speakers are genetically closer to Yoruba than they are to Bedouin, French or any non-African populations. Recent post-OOA non-African admixtures (bi-directional) through intermediaries explain why they are closer to Eurasians than Yoruba are to Eurasians. The Gabra population are still closer to Yoruba populations than to any Eurasians populations despite not having any evidence of recent admixture with Niger-Kordofanian speakers like Yoruba (who are also not recently admixed with any E1b1b populations).
To double check that Gabra from the Tiskoff study are also not recently admixed with NK speakers, we can also check Figure S12 in the Supporting Online Material posted before at K=14. We can see that Gabra have no orange color characteristic of Niger-Kordofanian speakers (orange at K=14 are mostly, but not only, recent NK alleles).
Double check done. Gabra are not recently admixed with Yoruba or any related NK populations. They have no e1b1a or L3e or orange color cluster. But they are still genetically closer to Yoruba and related Niger-Kordofanian speakers than to any Eurasian populations like Bedouin or French.
The reason why African populations like Cushitic speakers (eg. Gabra, Somali, etc) and NK speakers (eg Yoruba, Bantu, etc) are genetically closer to each others in relations to non-AFrican populations is because they share a common origin with other African populations from the same E-P2 Y-DNA lineage and various common MtDNA haplogroups lineages (L2a, L3bf, L3cd, L3eikx, L0a, etc).
So contrary to what some people try to say in this thread, we can see with both uniparental DNA haplogroups and autosomal DNA that Cushitic, Chadic and Niger-Kordofanian speakers share a common origin between the time of the OOA migrations of non-Africans and the foundation of Ancient Egypt.
Posted by Swenet (Member # 17303) on :
Quote: "Watch the clown cop-out again"
Exactly as anticipated. Notice the fact that this charlatan is his own source; his "evidence" tracks back to the spammy fairy tales he made up himself. If that doesn't ring alarm bells in the minds of the intelligent readers, I don't know what will.
quote:Originally posted by Swenet:
quote: Originally posted by Amun-Ra The Ultimate: For example, Yoruba got 92% of haplogroup P2/E1b1a, they share with East Africans like Somali, and only 8% of B-M60. This 8% represent what you call the aboriginal component to West Africans. Clearly it doesn't form the greater part of their ancestry.
This crappy statement presumes that haplogroup profiles are stable throughout time and accurate portrayals of all the genetic history of a population. Are they? Is E1b1a the only major Y-DNA expansion that impacted the ancestral Yoruba since West Africa was settled by AMHs? Post evidence.
^Watch the clown cop-out again
Posted by Tukuler (Member # 19944) on :
What I see above is ARtU presenting his argument and the methodology for his conclusions without resorting to ridicule, name-calling, baiting, etc.
Keep it up guy. Whereas this rationally makes your case, calling ESers racist did not.
BTW if you wanna counter what "some people" said why not confront them by replying to their post. I'm ready & waiting.
Posted by .Charlie Bass. (Member # 10328) on :
I wish the insults would stop for real. As far as I see, Pn2 clade not only unites different groups in Africa that speak different languages, it also links the same people to Europeans and Middle Eastern people who carry lineages under the same PN2 clade....paternally that is at some point in the past. All loci whether Y-Chromosones, mtDNA, X-Chromosone, or autosomal DNA, are important and do tell a story, it depends on what one is looking for, I wouldn't necessarily place one above the other because together analyzed you get a much bigger picture of what is going on.
Posted by Swenet (Member # 17303) on :
Y chromosomes and mtDNAs are the extremely sensitive to 101 agents of evolutionary change. They are absolutely worthless when it comes to unravelling the palimpsest layers of expansions that make each population what they are today, and even with the layers you are able to unravel by studying specific lineages, the frequencies in which they occur say absolutely nothing about the magnitude of the expansions that brought them to a particular locale.
Contrary to the fairy tales of these propagandists, 100% E1b1a doesn't mean the population considered originated with E1b1a. It doesn't even need to mean the majority of the genepool of that population originates with the ancestral population in which E1b1a emerged.
There are various genetic investigations one would need to reference to build a case for this bizarre claim, however, the charlatans deliberately avoid consulting these tests, because the result of said tests is the YRI population coalesces with other branches of Africans in the Middle Palaeolithic. Nowhere even near the terminal pleistocene touted as the magical moment West/Central Africans separated into their own biological entity.
Posted by .Charlie Bass. (Member # 10328) on :
quote:Originally posted by Swenet: Y chromosomes and mtDNAs are the extremely sensitive to 101 agents of evolutionary change. They are absolutely worthless when it comes to unravelling the palimpsest layers of expansions that make each population what they are today, and even with the layers you are able to unravel by studying specific lineages, the frequencies in which they occur say absolutely nothing about the magnitude of the expansions that brought them to a particular locale.
Contrary to the fairy tales of these propagandists, 100% E1b1a doesn't mean the population considered originated with E1b1a. It doesn't even need to mean the majority of the genepool of that population originates with the ancestral population in which E1b1a emerged.
There are various genetic investigations one would need to reference to build a case for this bizarre claim, however, the charlatans deliberately avoid consulting these tests, because the result of said tests is the YRI population coalesces with other branches of Africans in the Middle Palaeolithic. Nowhere even near the terminal pleistocene touted as the magical moment West/Central Africans separated into their own biological entity.
Man did you read what I said? Autosomals don't tell teh entire story either, you cannot look at autosomal ancestry and know someones Y-haplogroup or mtDNA, and the uni-parentals are good for tracking migrations from one region or another
Posted by Swenet (Member # 17303) on :
@Charlie
Of course I read what you said. While no one ever said that autosomes tell the whole story (a position you're bent on arguing against, but why?), someone did actually imply that haplogroup frequencies are more informative for uncovering all layers of migrations and their relative importance in terms of how much they're representative of the genetic make up of a population. I haven't seen you reply to that, once, hence, my previous attempt to put things back on track.
You're going out of your way to defend the usefullness of haplogroups, when, to my awareness, their value in general or ability to do what you're suggesting was never questioned. What exactly is it that you're arguing against, that makes you so determined to repeat your argument and feel like you're not being heard?
Quote: "Man did you read what I said?"
Posted by Djehuti (Member # 6698) on :
The point of the matter is that there are certain genetic markers only tell one side of the story. Uniparentals tell you exactly that-- one-parent lineages; while autosomes give you a rough picture of how you relate to others in everything else i.e. HLA. Even when both genetic methods are used, it still doesn't tell you their precise history as to which individuals migrated to their present location and mated with which individuals at a certain time. The best that genetics can give are CLUES as to the history of a population. It's just like archaeology. Archaeology gives clues via the material culture. History is perhaps the most accurate but of course written history as we know it is relatively recent and even the earliest histories written are full of mythology.
As for the author of this thread, Amun-Ra is obviously confused as he is biased to the point of blindness. He accuses us of being racist due to his mistaken belief that we are somehow white-washing or 'de-Africanizing' certain populations. We are NOT! However, he thinks this is the case due to the fact that the genetic diversity in Africa shows some populations are more closely related to certain populations while more distantly related to others.
Even the idiotic chart he spams shows this:
LOL Posted by .Charlie Bass. (Member # 10328) on :
I think people are over-emphasizing autosomes over the other loci, which should not be the case and I think it has a lot to do with the rise of these DNA companies and these STRUCTURE studies, to the point where people are seeing uni-parentals as useless and that should never be the case.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
^^^It's the contrary. Y-DNA and MtDNA analysis to determine your ancestors are very popular with DNA ancestry companies. They don't use autosomal much even if it could be done too. Ideally all three (Y-DNA, MtDNA, Autosomal) would be done at the same time for the same price. Giving you a more complete picture as I did in this thread:
quote:Originally posted by .Charlie Bass.: I think people are over-emphasizing autosomes over the other loci, which should not be the case and I think it has a lot to do with the rise of these DNA companies and these STRUCTURE studies, to the point where people are seeing uni-parentals as useless and that should never be the case.
I dont think this is the case. The two do not overlap. One can be easily altered by drift. Here is a clear example. Please match the autosomal profile to this Uniparental Marker.
E1b1b1 H1a
Where in Africa and the new world is this found?
Posted by .Charlie Bass. (Member # 10328) on :
quote:Originally posted by beyoku:
quote:Originally posted by .Charlie Bass.: I think people are over-emphasizing autosomes over the other loci, which should not be the case and I think it has a lot to do with the rise of these DNA companies and these STRUCTURE studies, to the point where people are seeing uni-parentals as useless and that should never be the case.
I dont think this is the case. The two do not overlap. One can be easily altered by drift. Here is a clear example. Please match the autosomal profile to this Uniparental Marker.
E1b1b1 H1a
Where in Africa and the new world is this found?
I never said that, but in the case of autosomals....look at Henry Louis gates, both his paternal and maternal uni-parentals are European, autosomally he is 50% African, yet he isn't a mulatto because neither of his parents are white, both are Aframs.
As for your question, well those two uniparentals you posted look about typical for a North African or even a Tuareg
Posted by beyoku (Member # 14524) on :
quote:Originally posted by .Charlie Bass.:
quote:Originally posted by beyoku:
quote:Originally posted by .Charlie Bass.: I think people are over-emphasizing autosomes over the other loci, which should not be the case and I think it has a lot to do with the rise of these DNA companies and these STRUCTURE studies, to the point where people are seeing uni-parentals as useless and that should never be the case.
I dont think this is the case. The two do not overlap. One can be easily altered by drift. Here is a clear example. Please match the autosomal profile to this Uniparental Marker.
E1b1b1 H1a
Where in Africa and the new world is this found?
I never said that, but in the case of autosomals....look at Henry Louis gates, both his paternal and maternal uni-parentals are European, autosomally he is 50% African, yet he isn't a mulatto because neither of his parents are white, both are Aframs.
As for your question, well those two uniparentals you posted look about typical for a North African or even a Tuareg
Or
That would have a totally different autosomal makeup than North Africans.....and would put the uniparental markers in perspective.
Posted by Swenet (Member # 17303) on :
Good point Beyoku. Hadn't even though of that in relation to this topic. Individuals with the same hgs but who belong to different populations will have an entirely different autosomal signature. If this happens at the population level (as opposed to the individual level), you get the very thing that Amun Ra's inner RAM memory can't seem to process without short circuiting: at the population level, individuals will cluster with the populations they're from, every time, regardless of how much the uniparental profiles of individuals within those populations resemble, or are identical to individuals from other, traditionally separated populations.
By far, in most population affinity uses, hgs are little more than icing on the cake or even third wheels compared to autosomal profiles. Sometimes they have specific niche benefits but the case can be made that many of these are either not population affinity/genetic closeness related (e.g. constructing trees that reflect the extant history of the human tree, entirely free of recombination) or that autosomes can perform some of those functions as well.
Posted by beyoku (Member # 14524) on :
This Reminds me of a discussion with a Somali. One of those nationalist "we not black....we not negro" types. I then proceeded to have him describe the Somali genesis and who was on Somalia prior to Somali and what negroids they have mixed with. He did a pretty bad job and tried to pin it all down to E-m78(v32). Somali are dominated by v32 but this marker only describes a paternal event in the area. It's hypothesized To have happened quite recent per Sanchez. ..I think 5600 or 6500 years ago.
Point is there are remains in Somalia that predate this. There is rock art in Somalia that dates to 6500 BC or earlier. It would be foolish to think Somali popped into existence with the dating of their main paternal marker. This marker is associated with s SMALL migration and has arisen to high frequency through drift.
Futuremore it is likely western Sudanese with high V32 have a different autosomal profile than Somali. According to limited STR at least they do.
Posted by beyoku (Member # 14524) on :
To add. What this parental marker DOES do, is let you get a picture of WHERE the small migration population came from before it rose to near fixation. It adds another element to the discussion. E-M81 reaching fixation in the Maghreb can be an anomaly when dealing with their autosomal profile....it does though let you know the Source of the founder in reg to the "founder effect".
The discussion in genetics has moved from uniparentals to Autosomal SNP's complimented by uniparentals. Many populations SHARE uniparentals. This is particularly the case with the area from Southern Europe through the fertile crescent and into the Horn and East Africa. If using the time span of "Dey all be E-PN2" as a gauge, then there is a vast span of lineages shared between Africans and non Africans when it comes to derivatives of M and N...Particularly RO. This would leads to vast swathes of that area mentioned above sharing R0 when not reduced all they way down to H. And Pn2. Folks in the chad basin would have an even closer sharing relationship with Europeans based on the sharing between Pn2 and R1.
The entire argument really falls flat when we look at NEW RESEARCH. Folks here dont know but haplogroup R and many lineages in south East Asia and the southern pacific have been consolidated and resolved down into SIMPLE SUBCLADES OF K2B. Yep you heard it....M, S, Q, R, P are all subcaldes of one K2b. If we take a step back to K2 we can include ALL of Haplogroup N and the VAST East Asian population of Haplogroup O.
http://en.wikipedia.org/wiki/Haplogroup_K-M9 The time frame of K2 breaking up in the diversity that it is today and all the populations it represents IS YOUNGER THAN the breakup of Haplogroup E and its subclades.
I will repeat that for emphasis and clarify:
The time frame of K2 breaking up in and Haplogroup diversity it represents today....and all the Autosomal separation between these populations (Europeans, Paupans, Chinese, Central Asians, Negritos, Southern Indians, Sub Saharans[T,R], Native Americans etc) IS YOUNGER THAN the breakup of Haplogroup E and its subclades....and possibly even E1b1 depending on the dating.
I know most of you dont study Eurasian genetics but to put things in perspective it would be very similar to finding new Y mutations in Africa that indicate that ALL the African uniparental markers can be resolved under a subclade of Sahelian E1a or East African B2a. once you understand the implications you can see how false it is to think Y dna is more important than or directly implies autosome.
Posted by Tukuler (Member # 19944) on :
are further from Fulani than Fulani are from European & Indian.
Cushitic are closer to to all Africans than they are to non-Africans, except * Hadza are further from Cushitic than Cushitic are from European & Indian.
Remember, the above are my observations of Tishkoff (2009r) Figure S14 aSTR nucleotide genetic distances measured tip to tip, not stem to stem.
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote:Originally posted by Tukuler:
Well, actually, the ordered tip to tip distance from European is
so, no, Fulani are not "closer to non-Africans than" are their Niger-Kordofanian co-linguists. The two are ~equidistant from European.
Well, actually, you're wrong.
And it's not the first time you fail to understand very basic things then come here to tell me, I'm the one who is wrong with an exaggerated confidence sure of yourself, instead of simply asking questions and ask me for clarification, if I'm able and willing to provide them. Obviously I can make simple errors too sometimes, all informed criticisms and counter-argumentation are welcome. It's a bit why I post those stuff here.
As I already explained to Beyoku (and thus to all readers of this forum) in greater length in another thread, that's not how you measure the distance between populations. You must not measure the distance from the tip but from the beginning of the colored lines.
If you check out p13-14 in the paragraph about Fig. S14 in the Tishkoff document ( DOWNLOAD HERE) . You will read:
The Fulani and Cushitic [] are closest to the non-African AACs. - Tiskhoff et al., P13-14
So it is indeed, as I implied in my previous post in this thread, Fulani and then Cushitic populations which are the closest to non-African AACs and the European AAC. Obviously, they are still much closer to the other African AACs than to the non-African ones.
As a side note, this is also reflected by the uniparental haplogroups of those populations which display a greater proportion of recent post-OOA Eurasian haplogroups admixtures (back migrations) in those populations.
Posted by Swenet (Member # 17303) on :
Why someone would rely on trees or dendrograms to infer pairwise genetic distances is completely beyond me.
Posted by .Charlie Bass. (Member # 10328) on :
quote:Originally posted by beyoku: To add. What this parental marker DOES do, is let you get a picture of WHERE the small migration population came from before it rose to near fixation. It adds another element to the discussion. E-M81 reaching fixation in the Maghreb can be an anomaly when dealing with their autosomal profile....it does though let you know the Source of the founder in reg to the "founder effect".
The discussion in genetics has moved from uniparentals to Autosomal SNP's complimented by uniparentals. Many populations SHARE uniparentals. This is particularly the case with the area from Southern Europe through the fertile crescent and into the Horn and East Africa. If using the time span of "Dey all be E-PN2" as a gauge, then there is a vast span of lineages shared between Africans and non Africans when it comes to derivatives of M and N...Particularly RO. This would leads to vast swathes of that area mentioned above sharing R0 when not reduced all they way down to H. And Pn2. Folks in the chad basin would have an even closer sharing relationship with Europeans based on the sharing between Pn2 and R1.
The entire argument really falls flat when we look at NEW RESEARCH. Folks here dont know but haplogroup R and many lineages in south East Asia and the southern pacific have been consolidated and resolved down into SIMPLE SUBCLADES OF K2B. Yep you heard it....M, S, Q, R, P are all subcaldes of one K2b. If we take a step back to K2 we can include ALL of Haplogroup N and the VAST East Asian population of Haplogroup O.
http://en.wikipedia.org/wiki/Haplogroup_K-M9 The time frame of K2 breaking up in the diversity that it is today and all the populations it represents IS YOUNGER THAN the breakup of Haplogroup E and its subclades.
I will repeat that for emphasis and clarify:
The time frame of K2 breaking up in and Haplogroup diversity it represents today....and all the Autosomal separation between these populations (Europeans, Paupans, Chinese, Central Asians, Negritos, Southern Indians, Sub Saharans[T,R], Native Americans etc) IS YOUNGER THAN the breakup of Haplogroup E and its subclades....and possibly even E1b1 depending on the dating.
I know most of you dont study Eurasian genetics but to put things in perspective it would be very similar to finding new Y mutations in Africa that indicate that ALL the African uniparental markers can be resolved under a subclade of Sahelian E1a or East African B2a. once you understand the implications you can see how false it is to think Y dna is more important than or directly implies autosome.
I agree with you bro, but the biggest flaw in regards to autosomals is when they try to use autosomals to track ancient population movements which to me in highly problematic. First reason is their proposed spread of autosomal ancestry is often at odds with the known inroduction and spread of certain mtDNA and Y haplogroups. I look at the case of the Khoisan, a couple of papers claim these people have anceint Eurasian admixture from East Africans, but there is no recognisable signature of anything Eurasian as far as haplogroups on neither the maternal nor paternal side.
The other thing is and we are seeing this is Europeans vs ancient European DNA being analysed, the autosomal make up of the ancients vs the moderns are very different and that can change several times over thousands of years. The same can happen for uniparentals but if there is a change you can track much more precisely on the uniparental side, thats how we know that thousands of years ago haplogroup A and B carriers(along with some now extinct lineages) were replaced by haplogroup E carriers, autosomals aren't as reliable in that regard. Autosomals are good for looking at the total genetic make up of populations if you are looking for percentages, but to me and this is jst me, they say nothing about ancient population movements. Look at that study on North African that proposed several movements of Eurasians into North Africa over 40,000 years
Posted by Amun-Ra The Ultimate (Member # 20039) on :
quote:Originally posted by .Charlie Bass.: I look at the case of the Khoisan, a couple of papers claim these people have anceint Eurasian admixture from East Africans, but there is no recognisable signature of anything Eurasian as far as haplogroups on neither the maternal nor paternal side.
That's not right. The percentage of Eurasian admixtures in Khoisan populations found in those autosomal studies is usually very low. But you can still see recognizable signature of Eurasian haplogroups in Khoisan populations. If we look at Y chromosome haplogroup frequencies table 3 on page 8 of this study ( download study here) Khoisan got 0.5% of Eurasian haplogroup I, 1.1% of R1a1 and 1.6% of R1b, all signatures of Eurasian haplogroups admixtures (post-OOA).
For a population like the Nama it's even higher (see here, fig3b p44) , in accordance to Pickrell (see here, Table 1) higher Eurasian autosomal finding for that particular population.
In general, you must always consider and take into account sample bias, SNP ascertainment bias and small sample size in those studies as I explained to you in another thread, as well as taking into account both uniparental DNA haplogroups and autosomal DNA as demonstrated in this thread.
Posted by beyoku (Member # 14524) on :
I think you misunderstood C-Bass.
Southern Africans were hypothesized to have "Eurasian" admixture via East Africans. East Africans have "Eurasians" admixture via southwest Asians. East Africans have the signatures of the admixture via uniparental markers: Generically speaking M89 and mtdna N.
The admixture in question goes back to a date in Horners where one would expect the Uniparental signature (M89 and mtdna N.) to be passed from East Africans to Southern Africans but that is not the case. Southern Africans instead only have signatures of Horn Ancestry...........indicating the South West Asian lineages in question were lost through drift or were never present in the Horn Africans that brought them.
Of course this does NOT take into account the very very recent EUROPEAN ancestry found in Southern Africans via rape and colonization of the area in the past few 100 years.
It is obvious that the research in question is looking at the Nile Valley/Rift valley under a lense that is too simplistic. Anyone that looks at the region should understand that there were a number of distinct groups in the Horn with differing uniparental profiles. The group...specifically the southern Cushitic speakers that traveled south and were absorbed into click speakers were distinct from other Horn populations affected by SW Asians. Off top I can think of at least 4 differing groups of East Africans dominated by certain uniparental markers that neighboring groups may not have had and have. All one needs to do is look at the discontinuity of lineages like V32, E-V6, L4, M1a etc. One southward expansion from the horn hypothetically would be of a population totally lacking in M78 while the other would not. The same could be said for M-329 and M-293.
IMO The horners in question that transverse to South Africa were absent M89. The M89 lineages found in Some horners is in fact more recent African mediated gene flow between neighboring populations. I believe it was Pickrell or that similar dated study on south Africans, when one looks at the expansion dates and admixture of all world populations Ethiopians have the oldest or second oldest expansion dates in ALL human groups sampled.
Posted by Swenet (Member # 17303) on :
The "Eurasian" population that admixed with Khoisans are projected to have been ~75% African. I don't see why that would have to leave a Eurasian hg signature in the recipient populations. I'm not saying there is no such hg signature, I'm saying that, in principle, there doesn't need to be one today. The very fact that select South Africans have notable freqs of hgs like L4, L5 and E-M293 today, and that these are especially found in South African pastoralists, proves that East Africans migrated there recently. This absence of substantial derived Eurasian hgs in these admixed Khoisan populations is clearly a(nother) case where hgs have proven unreliable trackers of migrations.
It was the study of autosomal DNA that brought clarity to the hitch-hiking layer of Eurasian ancestry towards South Africa, when hg investigations initially failed to attract researchers' undivided attention towards these "new" genetic aspects of the E-M293 migration, which go together well with what was initially known as movement of East African hgs, pottery and cattle to South Africa.
Posted by .Charlie Bass. (Member # 10328) on :
quote:Originally posted by Swenet: The "Eurasian" population that admixed with Khoisans are projected to have been ~75% African. I don't see why that would have to leave a Eurasian hg signature in the recipient populations. I'm not saying there is no such hg signature, I'm saying that, in principle, there doesn't need to be one today. The very fact that select South Africans have notable freqs of hgs like L4, L5 and E-M293 today, and that these are especially found in South African pastoralists, proves that East Africans migrated there recently. This absence of substantial derived Eurasian hgs in these admixed Khoisan populations is clearly a(nother) case where hgs have proven unreliable trackers of migrations.
It was the study of autosomal DNA that brought clarity to the hitch-hiking layer of Eurasian ancestry towards South Africa, when hg investigations initially failed to attract researchers' undivided attention towards these "new" genetic aspects of the E-M293 migration, which go together well with what was initially known as movement of East African hgs, pottery and cattle to South Africa.
I believe they see mixture and are trying to find a satisfactory date, thus all that talk about Aksum when in reality Aksum was a kingdom of its own without any significant mixture from the Red Sea. I think that specific "Eurasian2 ancestry is just Horn ancestry but of a different type
Posted by xyyman (Member # 13597) on :
@ Beyoku. Nice lead on K-M9. I, also, havent paid much attention to East Asian lineage until discovering N1c in equatorial West Africans. I just started getting into it.
Quote:
The entire argument really falls flat when we look at NEW RESEARCH. Folks here dont know but haplogroup R and many lineages in south East Asia and the southern pacific have been consolidated and resolved down into SIMPLE SUBCLADES OF K2B. Yep you heard it....M, S, Q, R, P are all subcaldes of one K2b. If we take a step back to K2 we can include ALL of Haplogroup N and the VAST East Asian population of Haplogroup O.
http://en.wikipedia.org/wiki/Haplogroup_K-M9 The time frame of K2 breaking up in the diversity that it is today and all the populations it represents IS YOUNGER THAN the breakup of Haplogroup E and its subclades.
I will repeat that for emphasis and clarify:
The time frame of K2 breaking up in and Haplogroup diversity it represents today....and all the Autosomal separation between these populations (Europeans, Paupans, Chinese, Central Asians, Negritos, Southern Indians, Sub Saharans[T,R], Native Americans etc) IS YOUNGER THAN the breakup of Haplogroup E and its subclades....and possibly even E1b1 depending on the dating
Posted by xyyman (Member # 13597) on :
Do you have the study?
Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome.- van Oven
Posted by xyyman (Member # 13597) on :
Havent paid much attention to this thread but what Bass said got me attention. I agree with Bass we needed to look at things holistically. HG completement AIM..
Also - to Beyoku's comment. Is the younger age of K2 indicative of a truly very recent OOA if AMH.
Some of us get confused with the letters and position in the Alphabet. The age of the HG is a lot more informative. Infact they need to adjust the nomenclature of the male and female PhyloTree
Posted by beyoku (Member # 14524) on :
^ how recent is recent?
Posted by xyyman (Member # 13597) on :
Do you have the study? It is behind the paywall. It is fascinating that K2 is now parental to these "Asian" lineage even R. Yet T, L R is found within Africa.
Keep in mind C6 is being found from Iberia to Siberia. lol! From 6000-45000 ya.
How recent is recent?
Posted by xyyman (Member # 13597) on :
Heh! you guys carry on. I just needed a copy or link of that study.
Posted by Swenet (Member # 17303) on :
quote:Originally posted by .Charlie Bass.:
quote:Originally posted by Swenet: The "Eurasian" population that admixed with Khoisans are projected to have been ~75% African. I don't see why that would have to leave a Eurasian hg signature in the recipient populations. I'm not saying there is no such hg signature, I'm saying that, in principle, there doesn't need to be one today. The very fact that select South Africans have notable freqs of hgs like L4, L5 and E-M293 today, and that these are especially found in South African pastoralists, proves that East Africans migrated there recently. This absence of substantial derived Eurasian hgs in these admixed Khoisan populations is clearly a(nother) case where hgs have proven unreliable trackers of migrations.
It was the study of autosomal DNA that brought clarity to the hitch-hiking layer of Eurasian ancestry towards South Africa, when hg investigations initially failed to attract researchers' undivided attention towards these "new" genetic aspects of the E-M293 migration, which go together well with what was initially known as movement of East African hgs, pottery and cattle to South Africa.
I believe they see mixture and are trying to find a satisfactory date, thus all that talk about Aksum when in reality Aksum was a kingdom of its own without any significant mixture from the Red Sea. I think that specific "Eurasian2 ancestry is just Horn ancestry but of a different type
I'm interested to see how you'd explain that there is a clear difference between the highly derived autosomal ancestry shared between East Africans and West Asians (i.e. the autosomal component that never seems to differentiate at higher Ks) and the non-derived autosomal ancestry shared between Ethiopians and West Asians (which does differentiate), and why, in the African continent, the former peaks in, and is mainly restricted to, Highland Ethiopians (i.e. occupants of the Axum area), while the latter is found throughout the wider region and seems to correspond to Tishkoff's "Cushitic" cluster.
If you ask me, the former autosomal component looks young and intrusive per all these specifics, and the fact that, among Highland Ethiopians, it's found more in speakers of a linguistic subclade that is also clearly intrusive and just as old as the immediate predecessors of the Axum kingdom, doesn't help.
Posted by .Charlie Bass. (Member # 10328) on :
Technically, autosomal mixture goes back as far as the mixture you get from your grandparents. I believe they are just seeing share components between some Eurasians and Horners, labelling it "Eurasian," then trying to fit it in some how.
Posted by beyoku (Member # 14524) on :
quote:Originally posted by .Charlie Bass.: Technically, autosomal mixture goes back as far as the mixture you get from your grandparents.
What? Did someone take control of your computer and post this?
Posted by Swenet (Member # 17303) on :
@Charlie
I get that that's your point, but why does the genetic material shared with Arabs that centers in the Ethiopian Highlands not resolve into it's on cluster at K=20, if it's old?
quote:Originally posted by .Charlie Bass.: Technically, autosomal mixture goes back as far as the mixture you get from your grandparents.
You'd have to explain that one to me..
Posted by .Charlie Bass. (Member # 10328) on :
What you just linked to speaks for itself. Beyoku also touched on this with the image that was discussed a page ago. The genetic material of all those ancestors is firmly lodged in our genome (no matter the size of the segments) so I don't see how that fits in the context in which you seemed to imply that the dating work done by Pagani and Pickrell is technically impossible and entirely the product of their own imagination and wishful thinking.
These are hefty accussations that you don't seem intent on backing up any time soon. You're obviously deliberately dancing around what you're asked, several times now, so, never mind.
Posted by .Charlie Bass. (Member # 10328) on :
Well take me for example. my mtDNA haplogroup is East African L4b2, no downstream, I have very minor east African ancestry but my mother and grandmother have it higher, get where I'm going? You can not reliably date autosomals and given recombination..........
Posted by Swenet (Member # 17303) on :
The estimates arrived at by the authors exploit recombination. So, not only do they work in spite of recombination; they leverage its effects on the genome.
The YRI - CEU dates reported for the controls (i.e. Aframs and N.Africans) in Pagani are on-point, as far as the most recent admixture events are concerned. Big vouch for the reliability of their estimates. The dates reported by Moorjani et al 2011, as far as the latest admixture events are concerned, are also on- point.
I can go on for a while. But I get it, you still think they're unreliable. Okay.
Posted by .Charlie Bass. (Member # 10328) on :
They are reliable but with limitations
Posted by Tukuler (Member # 19944) on :
quote:Originally posted by Swenet:
Why someone would rely on trees or dendrograms to infer pairwise genetic distances is completely beyond me.
.
I dunno??
Does Tishkoff (2009) present 5 "pairwise genetic distance trees or dendrograms" so that we should not rely on them to infer such distances?
Figure S14: Un-rooted neighbor-joining tree based on pairwise nucleotide genetic distances using inferred ancestral allele frequencies from the global STRUCTURE analysis at K = 14.
Figure S18: Un-rooted neighbor-joining tree from pairwise net nucleotide genetic distances calculated from the inferred ancestral allele frequencies at K = 14 from STRUCTURE analysis of the African dataset.
Figure S21: Un-rooted neighbor-joining tree from pairwise net nucleotide genetic distances calculated from the inferred ancestral allele frequencies for clusters at K = 10 with STRUCTURE analysis of the Eastern African dataset
I see no such tables, so, other than the above trees, where in this 2009 report does Tishkoff relate pairwise genetic information?
Posted by Tukuler (Member # 19944) on :
quote:Originally posted by Amun-Ra The Ultimate:
quote:Originally posted by Tukuler:
Well, actually, the ordered tip to tip distance from European is
so, no, Fulani are not "closer to non-Africans than" are their Niger-Kordofanian co-linguists. The two are ~equidistant from European.
Well, actually, you're wrong.
And it's not the first time you fail to understand very basic things then come here to tell me, I'm the one who is wrong with an exaggerated confidence sure of yourself, instead of simply asking questions and ask me for clarification, if I'm able and willing to provide them. Obviously I can make simple errors too sometimes, all informed criticisms and counter-argumentation are welcome. It's a bit why I post those stuff here.
As I already explained to Beyoku (and thus to all readers of this forum) in greater length in another thread, that's not how you measure the distance between populations. You must not measure the distance from the tip but from the beginning of the colored lines.
If you check out p13-14 in the paragraph about Fig. S14 in the Tishkoff document ( DOWNLOAD HERE) . You will read:
The Fulani and Cushitic [] are closest to the non-African AACs. - Tiskhoff et al., P13-14
So it is indeed, as I implied in my previous post in this thread, Fulani and then Cushitic populations which are the closest to non-African AACs and the European AAC. Obviously, they are still much closer to the other African AACs than to the non-African ones.
As a side note, this is also reflected by the uniparental haplogroups of those populations which display a greater proportion of recent post-OOA Eurasian haplogroups admixtures (back migrations) in those populations.
.
I didn't come here to tell you you were wrong. I came here to add my own observations about that NJ Tree.
You are absolutely correct.
Tishkoff's comments on Fig S14 are surely based on stem to stem measure not my mistaken tip to tip method which, however, is standard as Beyoku and the Lioness relayed from professional sources starting with your comment here followed by further discussion ending here.
I do not agree the colored bars represent, in your words, "within population genetic distance/"diversity" (not nucleotide diversity)" because Fig S14's label clearly states the tree represents nucleotide genetic distances whether between or within populations.
I am here to learn. Who are my teachers? All of you. Thanks for the correction ARtU. Will tuck it away in my "how-to-read" notes.

![[Big Grin]](biggrin.gif)
![[Roll Eyes]](rolleyes.gif)